Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Особенности организации генома эукариот. Классификации нуклеотидных последовательностей: уникальные, среднеповторяющиеся, высокоповторяющиеся. Регуляция экспрессии генов у эукариот.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Геном- совокупность генов гаплоидного набора хромосом. Ген- это линейный участок молекул ДНК, в котором закодирована последовательность а/к одной полипептидной цепи молекул белка. Большинство генов эукариот имеют мозаичное строение, т. е. состоят и чередующихся кодирующих (экзоны) и не кодирующих (интроны) участков. Последовательности: · Уникальные, т.е. последовательности, представленные в одном экземпляре или немногими копиями.
· Среднеповторяющиеся – последовательности, повторяющиеся сотни и тысячи раз.
· Высокоповторяющиеся, число которых достигает 10 миллионов на геном.
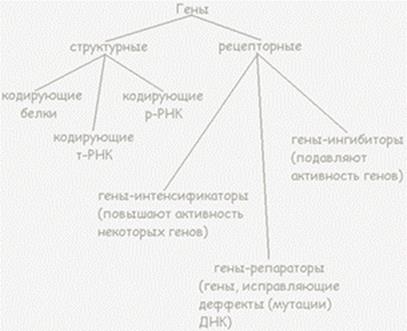
Регуляция экспрессии генов позволяет клеткам контролировать собственную структуру и функцию и является основой дифференцировки клеток, морфогенеза и адаптации. Экспрессия генов является субстратом для эволюционных изменений, так как контроль над временем, местом и количественными характеристиками экспрессии одного гена может иметь влияние на функции других генов в целом организме. Полученные к настоящему времени данные свидетельствуют, что экспрессия генов эукариот регулируется на уровне транскрипции, процессинга РНК в ядре и стабильности мРНК. Кроме того, было показано, что на экспрессию эукариотических генов оказывают влияние амплификация и перестройка генов. Классификация генов: структурные, функциональные (гены-модуляторы, ингибиторы, интенсификаторы, модификаторы); гены, регулирующие работу структурных генов (регуляторы и операторы), их роль в реализации наследственной информации. Структурные гены - это гены, контролирующие развитие конкретных признаков. Продуктом первичной активности гена является либо иРНК и далее полипептид, либо рРНК и тРНК. Таким образом, структурные гены содержат информацию об аминокислотных и нуклеотидных последовательностях макромолекул. При их мутациях наблюдаются обширные и разнообразные нарушения организма. Они образованы СРЕДЕПОВТОРЯЮЩИМИСЯ последовательностями ДНК. Виды структурных генов: · Функциональные гены, которые, в свою очередь, делятся на гены-модуляторы, усиливающие или ослабляющие действие структурных генов (ингибиторы, интенсификаторы, модификаторы) и гены, регулирующие работу структурных генов (регуляторы и операторы);
· Кодирующие аминокислотные последовательности структурных (коллаген) и ферментативных белков; · Кодирующие аминокислотные последовательности белков, функционирующих во всех клетках (например, рибосомных, гистонов); · Кодирующие последовательность нуклеотидов в молекулах рРНК и тРНК.
10) Множественные аллели как результат изменения нуклеотидной последовательности гена. Полиморфизм гена как вариант нормы и патологии. Примеры. Множественные аллели — один из видов взаимодействия аллельных генов, при котором ген может быть представлен не двумя аллелями (как в случаях полного или неполного доминирования), а гораздо большим их числом. Большинство генов существует в виде двух вариантов аллелей, но некоторые гены существуют в виде большого числа аллелей. Возникают они в результате многократных генных мутаций одного локуса гена. Чем больше аллельных генов, тем больше их комбинация попарно. У одной диплоидной особи может быть только два аллеля из серии множественных аллелей. Примеры: 1.множественные аллели окраски кроликов. Аллель С обеспечивает черную окраску тела; аллель ch - так называемую гималайскую окраску, когда черный цвет имеют уши, кончик морды, кончики лап и хвост; аллель с вызывает альбинизм. Аллель С доминирует над двумя другими, а аллель ch - над аллелем. 2. Наследование групп крови.
Полиморфными принято называть гены, которые представлены в популяции несколькими разновидностями - аллелями, что обусловливает разнообразие признаков внутри вида.
Обычно, причиной различий (полиморфизма) генов являются изменения отдельных нуклеотидов в молекуле ДНК, что приводит к изменению свойств гена (иногда в лучшую, а чаще, в худшую сторону). Некоторые изменения неизбежно являются причиной генных болезней и проявляются уже с рождения (например, муковисцидоз, мышечная дистрофия и др.), это – так называемые моногенные болезни, другие не приводят к болезням, но являются фактором предрасположенности к определенным заболеваниям (злокачественные опухоли, сердечно-сосудистые, аллергические и др. заболевания). В этом случае для развития болезни необходимы определенные внешние условия – характер питания, поступление в организм токсинов и онкогенов (табачный дым, алкоголь), нехватка витаминов и др. Эти болезни называются мультифакториальными. При определенных условиях (требуется достаточно длительный отрезок времени – сотни или тысячи лет) мутантные гены могут распространяться в популяциях и становиться достаточно обычными аллельными вариантами, обеспечивая основу генного полиморфизма.
Ген, его свойства. Примеры. Ген – элементарная функциональная единица наследственног материала, определяющая возможность развития отдельного признака клетки или организма. Свойства гена: · дискретность — несмешиваемость генов; · стабильность — способность сохранять структуру; · лабильность — способность многократно мутировать; · множественный аллелизм — многие гены существуют впопуляции во множестве молекулярных форм; · аллельность — в генотипе диплоидных организмов только двеформы гена; · специфичность — каждый ген кодирует свой признак; · плейотропия — множественный эффект гена; · экспрессивность — степень выраженности гена в признаке; · пенетрантность — частота проявления гена в фенотипе; · амплификация — увеличение количества копий гена. 12) Фенотип как результат реализации генотипа в конкретных условиях среды. Среда первого, второго (а и б) и третьего порядка. Экспрессивность и пенетрантность гена. Фенотип – совокупность биологических свойств и признаков организма, сложившаяся в процессе его индивидуального развития. Фенотипическое проявление генотипа в зависимости от среды изменяется в пределах нормы реакции. Норма реакции — пределы, в которых в зависимости от условий внешней среды может изменяться фенотипическое проявление отдельных генов или генотипа в целом. Совокупность внутриорганизменных факторов, влияющих на реализацию наследственной программы, обозначают как среду 1-го порядка. Особенно большое влияние на функцию генотипа факторы этой среды оказывают в период активных формообразовательных процессов, прежде всего в эмбриогенезе. С другой стороны, выделяют понятие окружающей среды, или среды 2-го порядка, как совокупности внешних по отношению к организму факторов. Степень проявления фенотипа – экспрессивность. Образно ее можно сравнить со степенью тяжести болезни в клинической практике. В основе изменчивости экспрессивности лежат и генетические факторы, и факторы внешней среды. Экспрессивность – очень важный показатель фенотипического проявления гена. Количественно ее степень определяют, используя статистический показатель. Генетический признак может даже не проявляться в некоторых случаях. Если ген есть в генотипе, но он вовсе не проявляется – он пенетрирован. Пенетрантность – количество особей (%), проявляющих в фенотипе данный ген, по отношению к количеству особей, у которых этот признак мог бы проявиться. Пенетрантность свойственна проявлению многих генов. Важен принцип – «все или ничего» - либо проявляется, любо нет. !!! Просмотреть задачи в конце второй рабочей тетради по биологии – полезно к экзамену.
13) Определение пола. Первичные и вторичные половые признаки. Половой диморфизм. Типы определения пола: прогамный, сингамный, эпигамный. Хромосомный механизм определения пола у разных организмов. Гомогаметный и гетерогаметный пол. Роль генотипа и среды в развитии признаков пола.
Пол у большинства животных и растений определяется генетически в момент оплодотворения. При исследовании кариотипов многих животных было установлено,что у женского организма каждая хромосома имеет парную (идентичную по размерам,морфологии и содержанию генов),а у мужских организмов имеются две непарные хромосомы, которые резко отличаются по величине, морфологии и заключенной в них генетической информации. При дальнейшем исследовании было показано,что эти непарные хромосомы и определяют пол огранизма. Их назвали половыми хромосомами, в отличие от других – аутосом.Большую из непарных хромосом, одинаковую у мужского и женского организмов, назвали Х-хромосомой, а меньшую, имеющуюся только у мужского организма - Y-хромосомой. Половые признаки — совокупность признаков, по которым мужской и женский пол отличают друг от друга. Половые признаки бывают первичными, или основными, и вторичными. Первые — это половые железы — яичники или семенники. Вторичные половые признаки формируются в течение роста и полового созревания организма. У мужчин они проявляются в росте бороды, усов, появлении низкого тембра голоса и др., у женщин — в развитии молочных желез, в появлении определенных особенностей телосложения и других признаков. У человека и позвоночных животных вторичные половые признаки являются функцией деятельности половых желез. Интенсивность полового созревания людей зависит от социально-бытовых условий, наследственности и других причин. Половой диморфизм — анатомические различия между самцами и самками одного и того же биологического вида, исключая различия в строении половых органов. Половой диморфизм может проявляться в различных физических признаках:
· Размер. У большинства млекопитающих и многих видов птиц самцы более крупные и тяжёлые, чем самки. У земноводных и членистоногих самки, как правило, крупнее самцов. У черепах и у змей самки крупнее самцов; у ящериц, напротив, как правило, самцы крупнее самок; исключение составляют гекконы, у которых большая величина также свойственна самкам. · Волосяной покров. Борода у мужчин, грива у львов или бабуинов. · Окраска. Цвет оперения у птиц, особенно у утиных. · Кожа. Характерные наросты или дополнительные образования, такие как рога у оленевых, гребешок у петухов. · Зубы. Бивни у самцов индийского слона, более крупные клыки у самцов моржей и кабанов. Типы определения пола: · Прогамный; прогамное определение пола происходит до оплодотворения, в процессе формирования яйцеклеток. Это имеет место, например, у коловраток. Они образуют яйцеклетки двух сортов: крупные, с двумя наборами хромосом (диплоидные) и большим объёмом цитоплазмы и мелкие, с одним набором хромосом — гаплоидные. Из гаплоидных неоплодотворённых яиц развиваются гаплоидные самцы, продуцирующие гаплоидные гаметы. Из крупных диплоидных яиц также развиваются самки, но в этом случае они появляются не в результате оплодотворения, а партеногенетически, то есть без оплодотворения. Таким образом, пол особи, развивающейся из диплоидного яйца, определяется ещё на стадии формирования яйцеклетки (на этом этапе закладывается её диплоидность), а пол особи, развивающейся из мелкого яйца, зависит от того, будет оно оплодотворено или нет;
· Эпигамный; под влиянием внешней среды – например, температуры; У некоторых рептилий пол определяется температурой, в которой развивалось яйцо в течение температурочувствительного периода. Это явление носит название температурозависимого определения пола; · Сингамный; сингамное определение пола происходит при оплодотворении, и пол определяется генетически. Хромосомный механизм определения пола у разных организмов
У большинства организмов женский пол гомогаметный (хх),а мужской – гетерогаметный (ху). У птиц, бабочек, пресмыкающихся, гомогаметен (хх) мужской пол. У многих насекомых, например, у кузнечика, у некоторых моллюсков, у кенгуру Y-хромосомы вообще нет. У них клетки женских особей содержат две Х хромосомы, а клетки мужской - одну Х-хромосому (хо). У млекопитающих развитие половых признаков мужского пола обусловлено значительным количеством генов, большая часть которых расположена в аутосомах. Но начинают они работать только при наличии фактора, который расположен на маленькой Y-хромосоме. Развитие зародышей крокодила у мужских и женских особей определяется температурой, при которой происходит инкубация яиц.32-33 градуса - самцы, выше или ниже - самки. Развитие пола морского беспозвоночного бонелии определяется влиянием внешних факторов. Этот фактор – наличие взрослой самки. Если ее нет, то личинки развиваются только как самки. Если есть - только как самцы. Хромосомная теория наследственности Т. Моргана. Основные положения. Группы сцепления. Группы сцепления у гомо- и гетерогаметного пола у человека. “Цис-“, “транс-“ положение генов в группах сцепления. Кроссинговер как механизм, нарушающий сцепление генов. Построение хромосомных карт. Хромосомная теория наследственности — теория, согласно которой передача наследственной информации в ряду поколений связана с передачей хромосом, в которых в определённой и линейной последовательности расположены гены. Анализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности: · Гены находятся в хромосомах. · Гены расположены в хромосоме в линейной последовательности.
· Различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален. · Аллельные гены занимают одинаковые локусы в гомологичных хромосомах. Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцепленно (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола). Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами). Каждый биологический вид характеризуется определенным набором хромосом — кариотипом.
Группа сцепления — совокупность генов, находящихся в одной хромосоме. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола). Кроссинговер, перекрест, взаимный обмен участками парных хромосом, происходящий в результате разрыва и соединения в новом порядке их нитей — хроматид; приводит к перераспределению (рекомбинации) сцепленных генов. Важнейший механизм, обеспечивающий комбинаторную изменчивость, а следовательно, — один из главных факторов эволюции. К., как правило, имеет место в профазе первого деления половых клеток, когда их хромосомы представлены четырьмя нитями. Генети́ческая ка́рта — схема взаимного расположения структурных генов, регуляторных элементов игенетических маркеров, а также относительных расстояний между ними на хромосоме. Генетические карты составляют для каждой пары гомологичных хромосом. Каждой паре присваивается номер (I, II, III и т.д.), группы сцепления номеруются в порядке их обнаружения. Кроме номера в каждой из групп сцепления указывают полное или сокращенное название генов, расстояние этих генов в единицах перекреста от одного из концов хромосомы, а также место расположения центромеры. Следует отметить, что длина хромосомы не обязательно является показателем ее генетической активности. Для генетических карт применяется термин «локус» для обозначения места гена в хромосоме или на ее карте. Пример записи цис- и транс- положения: Цис-положение - расположение тесно сцепленных рецессивных аллелей двух или нескольких локусов в одной из хромосом гомологичной пары, а доминантных — в другой. Транс-положение - локализация 2 данных аллелей (генов) на разных гомологичных хромосомах.
Цитоплазматическая наследственность. Плазмогены. Наследование внеядерных генов. Закономерности наследования внеядерных генов. Плазмиды; их роль в передаче наследственной информации у прокариот. Горизонтальный перенос генов. Наследственность цитоплазматическая (внеядерная, нехромосомная, плазматическая) - преемственность материальных структур и функциональных свойств организма, которые определяются и передаются факторами, расположенными в цитоплазме. Совокупность этих факторов - плазмогенов, или внеядерных генов, составляет плазмон (подобно тому, как совокупность хромосомных генов - геном). Плазмогены находятся в самовоспроизводящихся органеллах клетки - митохондриях и пластидах. Плазмогены передаются главным образом через женскую половую клетку (яйцеклетку), так как мужская половая клетка (спермий) почти не содержит цитоплазмы (что, однако, не исключает передачи плазмогенов через мужские гаметы). Поэтому изучение цитоплазматической наследственности ведётся с использованием специальных схем скрещивания, при которых данный организм (или группа) используется и как материнская, и как отцовская форма (реципрокное скрещивание). У растений и животных различия, обусловленные цитоплазматической наследственностью, сводятся в основном к преобладанию материнских признаков и проявлению определённого фенотипа при одном направлении скрещивания и его утрате при другом. Прокариоты, помимо генов, которые заключены в хромосомную ДНК, имеют небольшой внехромосомный набор генов, или по-другому – плазмиды. У некоторых плазмид есть свойство встраиваться в хромосомы хозяйских клеток, тогда такая разновидность пластид называется эписомами. Бактериальные плазмиды представляют собой двухцепочечные, небольшие по размерам кольцевидные молекулы ДНК, которые имеют способность удваиваться отдельно от хромосомы хозяина. Только плазмиды, которые входят в состав хромосом бактерий, удваиваются вместе с бактерией. В гены многих плазмидов заключена информация, которая в последующем будет влиять на фенотип клетки, в которой находится плазмида. Горизонтальный перенос генов (ГПГ) — процесс, в котором организм передаёт генетический материал другому организму, не являющемуся его потомком. В отличие от горизонтального, о вертикальном переносе генов говорят, если организм получает генетический материал от своего предка. В области интересов генетики основное место занимает вертикальный перенос генов. Однако в настоящее время горизонтальному переносу уделяется всё больше внимания. Репарация генетического материала. Фотореактивация. Темновая репарация. Этапы. Мутации, связанные с нарушением репарации. Примеры.
Главным антимутационным барьером рассматривается выработавшая в процессе эволюции способность к репарации наследственного материала. Её сущность - в устранении из наследственного материала клетки изменённого участка. Различают 3 системы репарации генетического материала: · эксцизионная репарация (репарация путём «вырезания»); · фоторепарация; · пострепликативная репарация; Эксцизионная репарация - удаление повреждённых азотистых оснований из ДНК и последующее восстановление нормальной структуры молекулы по комплементарной цепи. Ферментативная система удаляет короткую однонитевую последовательность двунитевой ДНК, содержащей ошибочно спаренные или поврежденные основания, и замещает их путём синтеза последовательности, комплементарной оставшейся нити. Фоторепарация (фотореактивация) - является наиболее простым механизмом репарации ДНК. Заключается в расщеплении ферментом (дезоксирибо-пиримидинфотолиазой), активируемым видимым светом, циклобутановых димеров, возникающих в ДНК под действием ультрафиолетового излучения. Пострепликативная репарация - пострепликативная репарация «включается» тогда, когда эксцизионная репарация «не справляется» с устранением всех повреждений, возникших в ДНК до её репликации. При репликации во второй спирали ДНК возникают бреши - однонитевые пробелы, соответствующие изменённым нуклеотидам первой спирали. Бреши заполняются участками цепи с нормальной последовательностью нуклеотидов уже в ходе пострешгикативной репарации при участии ДНК-полимеразы. Позднее была обнаружена и темновая репарация, т. е. свойство клеток ликвидировать повреждения ДНК без участия видимого света. Темновая репарация осуществляется комплексом из пяти ферментов:
· узнающего химические изменения на участке цепи ДНК; · осуществляющего вырезание поврежденного участка; · удаляющего этот участок; · синтезирующего новый участок по принципу комплементарности взамен удаленного фрагмента; · соединяющего концы старой цепи и восстановленного участка. При световой репарации исправляются повреждения, возникшие только под воздействием ультрафиолетовых лучей, при темновой — повреждения, появившиеся под влиянием жесткой радиации, химических веществ и других факторов. Темновая репарация обнаружена как у прокариот, так и в клетках эукариот. У последних она изучается в культурах тканей. Вопрос о том, почему одни повреждения репарируются, а другие нет, остается открытым. Если репарация не наступает, то клетка либо гибнет, либо наступает мутация.
|
|||||||||
|
|
Последнее изменение этой страницы: 2019-05-19; просмотров: 782; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.133.143.183 (0.014 с.) |