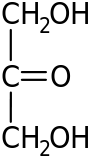Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Нуклеиновые кислоты. Строение, функции. Принцип матричного синтеза как основа наследственных свойств живых систем. Генетический кодСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Полимерные формы нуклеиновых кислот называют полинуклеотидами. Цепочки из нуклеотидов соединяются через остаток фосфорной кислоты(фосфодиэфирная связь). Поскольку в нуклеотидах существует только два типа гетероциклических молекул, рибоза и дезоксирибоза, то и имеется лишь два вида нуклеиновых кислот — дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК). Мономерные формы также встречаются в клетках и играют важную роль в процессах передачи сигналов или запасании энергии. Наиболее известный мономер РНК — АТФ, аденозинтрифосфорная кислота, важнейший аккумулятор энергии в клетке. Подобно белкам, нуклеиновые кислоты — биополимеры, а их функция заключается в хранении, реализации и передаче генетической (наследственной) информации в живых организмах. Существует два типа нуклеиновых кислот — дезоксирибонуклеиновые (ДНК) и рибонуклеиновые (РНК). Мономерами в нуклеиновых кислотах служат нуклеотиды. Каждый из них содержит азотистое основание, пятиуглеродный сахар (дезоксирибоза — в ДНК, рибоза — в РНК) и остаток фосфорной кислоты. В ДНК входят четыре вида нуклеотидов, отличающихся по азотистому основанию в их составе, — аденин (А), гуанин (Г), цитозин (Ц) и тимин (Т). В молекуле РНК также имеется 4 вида нуклеотидов с одним из азотистых оснований — аденином, гуанином, цитозином и урацилом (У). Таким образом, ДНК и РНК различаются как по содержанию сахара в нуклеотидах, так и по одному из азотистых оснований (табл. 1).
Компоненты нуклеотидов ДНК и РНК
Молекулы ДНК и РНК существенно различаются по своему строению и выполняемым функциям. Молекула ДНК может включать огромное количество нуклеотидов — от нескольких тысяч до сотен миллионов (поистине гигантские молекулы ДНК удается «увидеть» с помощью электронного микроскопа). В структурном отношении она представляет собой двойную спираль из полинуклеотидных цепей (рис. 1), соединенных с помощью водородных связей между азотистыми основаниями нуклеотидов. Благодаря этому полинуклеотидные цепи прочно удерживаются одна возле другой. При исследовании различных ДНК (у разных видов организмов) было установлено, что аденин одной цепи может связываться лишь с тимином, а гуанин — только с цитозином другой. Следовательно, порядок расположения нуклеотидов в одной цепи строго соответствует порядку их расположения в другой. Этот феномен получил название комплементарности (т. е. дополнения), а противоположные полинуклеотидные цепи называются комплементарными. Именно этим обусловлено уникальное среди всех неорганических и органических веществ свойство ДНК — способность к самовоспроизведению или удвоению (рис. 2). При этом сначала комплементарные цепи молекул ДНК расходятся (под воздействием специального фермента происходит разрушение связей между комплементарными нуклеотидами двух цепей). Затем на каждой цепи начинается синтез новой («недостающей») комплементарной ей цепи за счет свободных нуклеотидов, всегда имеющихся в большом количестве в клетке. В результате вместо одной («материнской») молекулы ДНК образуются две («дочерние») новые, идентичные по структуре и составу друг другу, а также исходной молекуле ДНК. Этот процесс всегда предшествует клеточному делению и обеспечивает передачу наследственной информации от материнской клетки дочерним и всем последующим поколениям. Молекулы РНК, как правило, одноцепочечные (в отличие от ДНК) и содержат значительно меньшее число нуклеотидов. Выделяют три вида РНК (табл. 2), различающиеся по величине молекул и выполняемым функциям, — информационную (иРНК), рибосомальную (рРНК) и транспортную (тРНК). Три вида РНК
Информационная РНК (и-РНК) располагается в ядре и цитоплазме клетки, имеет самую длинную полинуклеотидную цепь среди РНК и выполняет функцию переноса наследственной информации из ядра в цитоплазму клетки. Транспортная РНК (т-РНК) также содержится в ядре и цитоплазме клетки, ее цепь имеет наиболее сложную структуру, а также является самой короткой (75 нуклеотидов). Т-РНК доставляет аминокислоты к рибосомам в процессе трансляции — биосинтеза белка. Рибосомальная РНК (р-РНК) содержится в ядрышке и рибосомах клетки, имеет цепь средней длины. Все виды РНК образуются в процессе транскрипции соответствующих генов ДНК.
Углеводы, липиды. Функции, строение Липиды — один из важнейших классов сложных молекул, присутствующих в клетках и тканях животных. Липиды выполняют самые разнообразные функции: снабжают энергией клеточные процессы, формируют клеточные мембраны, участвуют в межклеточной и внутриклеточной сигнализации. Липиды служат предшественниками стероидных гормонов,желчных кислот, простагландинов и фосфоинозитидов. В крови содержатся отдельные компоненты липидов (насыщенные жирные кислоты, мононенасыщенные жирные кислоты и полиненасыщенные жирные кислоты), триглицериды, холестерин, эфиры холестерина и фосфолипиды. Все эти вещества не растворимы в воде, поэтому в организме имеется сложная система транспорта липидов. Свободные (неэтерифицированные) жирные кислоты переносятся кровью в виде комплексов с альбумином. Триглицериды, холестерин и фосфолипиды транспортируются в форме водорастворимых липопротеидов. Некоторые липиды используются для создания наночастиц, например, липосом. Мембрана липосом состоит из природных фосфолипидов, что определяет их многие привлекательные качества. Они нетоксичны, биодеградируемы, при определенных условиях могут поглощаться клетками, что приводит к внутриклеточной доставке их содержимого. Липосомы предназначены для целевой доставки в клетки препаратов фотодинамической или генной терапии, а также компонентов другого назначения, например, косметического[3]. Строение Молекулы простых липидов состоят из спирта, жирных кислот, сложные — из спирта, высокомолекулярных жирных кислот, возможны остатки фосфорной кислоты, углеводов, азотистых оснований и др. Строение липидов зависит в первую очередь от пути их биосинтеза. Биологические функции Энергетическая (резервная) функция Многие жиры, в первую очередь триглицериды, используются организмом как источник энергии. При полном окислении 1 г жира выделяется около 9 ккал энергии, примерно вдвое больше, чем при окислении 1 г углеводов (4.1 ккал). Жировые отложения используются в качестве запасных источников питательных веществ, прежде всего животными, которые вынуждены носить свои запасы на себе. Растения чаще запасают углеводы, однако в семенах многих растений высоко содержание жиров (растительные масла добывают из семян подсолнечника, кукурузы, рапса, льна и других масличных растений). Функция теплоизоляции Жир — хороший теплоизолятор, поэтому у многих теплокровных животных он откладывается в подкожной жировой ткани, уменьшая потери тепла. Особенно толстый подкожный жировой слой характерен для водных млекопитающих (китов, моржей и др.). Но в то же же время у животных, обитающих в условиях жаркого климата (верблюды, тушканчики) жировые запасы откладываются на изолированных участках тела (в горбах у верблюда, в хвосте у жирнохвостых тушканчиков), в качестве резервных запасов воды, так как вода — один из продуктов окисления жиров. Структурная функция Фосфолипиды составляют основу биослоя клеточных мембран, холестерин — регулятор текучести мембран. У архей в состав мембран входят производные изопреноидных углеводородов. Воски образуют кутикулу на поверхности надземных органов (листьев и молодых побегов) растений. Их также производят многие насекомые (так, пчёлы строят из них соты, а червецы и щитовки образуют защитные чехлы).
Регуляторная · Витамины — липиды (A, D, E, K) · Гормональная (стероиды, эйкозаноиды, простагландины и прочие.) · Кофакторы (долихол) · Сигнальные молекулы (диглицериды, жасмоновая кислота; МP3-каскад) Защитная (амортизационная) Толстый слой жира защищает внутренние органы многих животных от повреждений при ударах (например, сивучи при массе до тонны, могут прыгать в воду со скал высотой 20-25м) Увеличения плавучести Самые разные организмы — от диатомовых водорослей до акул — используют резервные запасы жира как средство снижения среднего удельного веса тела и, таким образом, увеличения плавучести. Это позволяет снизить расходы энергии на удержание в толще воды. Углеводы Простые и сложные Углеводы являются неотъемлемым компонентом клеток и тканей всех живых организмов представителей растительного и животного мира, составляя (по массе) основную часть органического вещества на Земле. Источником углеводов для всех живых организмов является процесс фотосинтеза, осуществляемый растениями. По способности к гидролизу на мономеры углеводы делятся на две группы: простые (моносахариды) и сложные (дисахариды и полисахариды). Сложные углеводы в отличие от простых способны гидролизоваться с образованием моносахаридов, мономеров. Простые углеводы легко растворяются в воде и синтезируются в зелёных растениях. Сложные углеводы являются продуктами поликонденсации простых сахаров (моносахаридов), а в процессе гидролитического расщепления образуют сотни и тысячи молекул моносахаридов[2].
Моносахариды
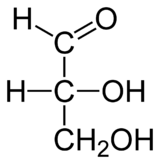
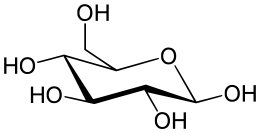
Распространённый в природе моносахарид — бета-D-глюкоза. Моносахари́ды (от греческого monos — единственный, sacchar — сахар) — простейшие углеводы, не гидролизующиеся с образованием более простых углеводов — обычно представляют собой бесцветные, легко растворимые в воде, плохо — в спирте и совсем нерастворимые в эфире, твёрдые прозрачные органические соединения[2], одна из основных групп углеводов, самая простая форма сахара. Водные растворы имеют нейтральную pH. Некоторые моносахариды обладают сладким вкусом. Моносахариды содержат карбонильную (альдегидную или кетонную) группу, поэтому их можно рассматривать как производные многоатомных спиртов. Моносахарид, у которого карбонильная группа расположена в конце цепи, представляет собой альдегид и называется альдоза. При любом другом положении карбонильной группы моносахарид является кетоном и называется кетоза. В зависимости от длины углеродной цепи (от трёх до десяти атомов) различают триозы, тетрозы, пентозы, гексозы, гептозы и так далее. Среди них наибольшее распространение в природе получили пентозы и гексозы[2]. Моносахариды — стандартные блоки, из которых синтезируются дисахариды, олигосахариды и полисахариды. В природе в свободном виде наиболее распространена D-глюкоза (виноградный сахар или декстроза, C 6 H 12 O 6) — шестиатомный сахар (гексоза), структурная единица (мономер) многих полисахаридов (полимеров) — дисахаридов: (мальтозы, сахарозы и лактозы) и полисахаридов (целлюлоза, крахмал). Другие моносахариды, в основном, известны как компоненты ди-, олиго- или полисахаридов и в свободном состоянии встречаются редко. Природные полисахариды служат основными источниками моносахаридов[2]. Дисахариды
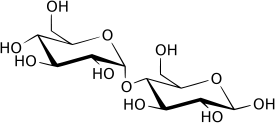
Мальтоза (солодовый сахар) — природныйдисахарид, состоящий из двух остатков глюкозы Дисахари́ды (от di — два, sacchar — сахар) — сложные органические соединения, одна из основных групп углеводов, при гидролизе каждая молекула распадается на две молекулы моносахаридов, являются частным случаем олигосахаридов. По строению дисахариды представляют собой гликозиды, в которых две молекулы моносахаридов соединены друг с другом гликозидной связью, образованной в результате взаимодействия гидроксильных групп (двух полуацетальных или одной полуацетальной и одной спиртовой). В зависимости от строения дисахариды делятся на две группы: восстанавливающие и невосстанавливающие. Например, в молекуле мальтозы у второго остатка моносахарида (глюкозы) имеется свободный полуацетальный гидроксил, придающий данному дисахариду восстанавливающие свойства. Дисахариды наряду с полисахаридами являются одним из основных источников углеводов в рационе человека и животных[3]. Олигосахариды
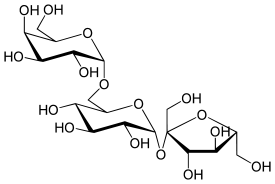
Рафиноза — природный трисахарид, состоящий из остатков D-галактозы, D-глюкозы и D-фруктозы. О́лигосахари́ды (от греч. ὀλίγος — немногий) — углеводы, молекулы которых синтезированы из 2 — 10 остатков моносахаридов, соединённых гликозидными связями. Соответственно различают: дисахариды, трисахариды и так далее[3]. Олигосахариды, состоящие из одинаковых моносахаридных остатков, называют гомополисахаридами, а из разных — гетерополисахаридами. Наиболее распространены среди олигосахаридов дисахариды. Среди природных трисахаридов наиболее распространена рафиноза — не восстанавливающий олигосахарид, содержащий остатки фруктозы, глюкозы и галактозы — в больших количествах содержится в сахарной свёкле и во многих других растениях[3]. Полисахариды
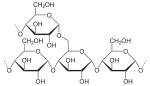
Полисахари́ды — общее название класса сложных высокомолекулярных углеводов, молекулы которых состоят из десятков, сотен или тысяч мономеров — моносахаридов. С точки зрения общих принципов строения в группе полисахаридов возможно различить гомополисахариды, синтезированные из однотипных моносахаридных единиц и гетерополисахариды, для которых характерно наличие двух или нескольких типов мономерных остатков[4]. Гомополисахариды (гликаны), состоящие из остатков одного моносахарида, могут быть гексозами или пентозами, то есть в качестве мономера может быть использована гексоза или пентоза. В зависимости от химической природы полисахарида различают глюканы (из остатков глюкозы), маннаны (из маннозы), галактаны (из галактозы) и другие подобные соединения. К группе гомополисахаридов относятся органические соединения растительного (крахмал,целлюлоза, пектиновые вещества), животного (гликоген, хитин) и бактериального (декстраны) происхождения[2]. Полисахариды необходимы для жизнедеятельности животных и растительных организмов. Это один из основных источников энергии организма, образующейся в результате обмена веществ. Полисахариды принимают участие в иммунных процессах, обеспечивают сцепление клеток в тканях, являются основной массой органического вещества в биосфере. Крахма́л (C 6 H 10 O 5)n — смесь двух гомополисахаридов: линейного — амилозы и разветвлённого — амилопектина, мономером которых является альфа-глюкоза. Белое аморфное вещество, не растворимое в холодной воде, способное к набуханию и частично растворимое в горячей воде[2]. Молекулярная масса 105—107 Дальтон. Крахмал, синтезируемый разными растениями в хлоропластах, под действием света при фотосинтезе, несколько различается по структуре зёрен, степени полимеризации молекул, строению полимерных цепей и физико-химическим свойствам. Как правило, содержание амилозы в крахмале составляет 10—30 %, амилопектина — 70—90 %. Молекула амилозы содержит в среднем около 1 000 остатков глюкозы, связанных между собой альфа-1,4-связями. Отдельные линейные участки молекулы амилопектина состоят из 20—30 таких единиц, а в точках ветвления амилопектина остатки глюкозы связаны межцепочечными альфа-1,6-связями. При частичном кислотном гидролизе крахмала образуются полисахариды меньшей степени полимеризации — декстрины (C 6 H 10 O 5)p, а при полном гидролизе — глюкоза[4]. Гликоге́н (C 6 H 10 O 5)n — полисахарид, построенный из остатков альфа-D-глюкозы — главный резервный полисахарид высших животных и человека, содержится в виде гранул вцитоплазме клеток практически во всех органах и тканях, однако, наибольшее его количество накапливается в мышцах и печени. Молекула гликогена построена из ветвящихся полиглюкозидных цепей, в линейной последовательности которых, остатки глюкозы соединены посредством альфа-1,4-связями, а в точках ветвления межцепочечными альфа-1,6-связями. Эмпирическая формула гликогена идентична формуле крахмала. По химическому строению гликоген близок к амилопектину с более выраженной разветвлённостью цепей, поэтому иногда называется неточным термином «животный крахмал». Молекулярная масса 105—108 Дальтон и выше[4]. В организмах животных является структурным и функциональным аналогом полисахарида растений — крахмала. Гликоген образует энергетический резерв, который при необходимости восполнить внезапный недостаток глюкозы может быть быстро мобилизован — сильное разветвление его молекулы ведёт к наличию большого числа концевых остатков, обеспечивающих возможность быстрого отщепления нужного количества молекул глюкозы[2]. В отличие от запаса триглицеридов (жиров) запас гликогена не настолько ёмок (в калориях на грамм). Только гликоген, запасённый в клеткахпечени (гепатоцитах) может быть переработан в глюкозу для питания всего организма, при этом гепатоциты способны накапливать до 8 процентов своего веса в виде гликогена, что является максимальной концентрацией среди всех видов клеток. Общая масса гликогена в печени взрослых может достигать 100—120 граммов. В мышцах гликоген расщепляется на глюкозу исключительно для локального потребления и накапливается в гораздо меньших концентрациях (не более 1 % от общей массы мышц), тем не менее общий запас в мышцах может превышать запас, накопленный в гепатоцитах. Целлюло́за (клетча́тка) — наиболее распространённый структурный полисахарид растительного мира, состоящий из остатков альфа-глюкозы, представленных в бета-пиранозной форме. Таким образом, в молекуле целлюлозы бета-глюкопиранозные мономерные единицы линейно соединены между собой бета-1,4-связями. При частичном гидролизе целлюлозы образуется дисахарид целлобиоза, а при полном — D-глюкоза. В желудочно-кишечном тракте человека целлюлоза не переваривается, так как набор пищеварительных ферментов не содержит бета-глюкозидазу. Тем не менее, наличие оптимального количества растительной клетчатки в пище способствует нормальному формированию каловых масс[4]. Обладая большой механической прочностью, целлюлоза выполняет роль опорного материала растений, например, в составе древесины её доля варьирует от 50 до 70 %, а хлопок представляет собой практически стопроцентную целлюлозу[2]. Хити́н — структурный полисахарид низших растений, грибов и беспозвоночных животных (в основном роговые оболочки членистоногих — насекомых и ракообразных). Хитин, подобно целлюлозе в растениях, выполняет опорные и механические функции в организмах грибов и животных. Молекула хитина построена из остатков N-ацетил-D-глюкозамина, связанных между собой бета-1,4-гликозиюными связями. Макромолекулы хитина неразветвлённые и их пространственная укладка не имеет ничего общего с целлюлозой[2]. Пекти́новые вещества́ — полигалактуроновая кислота, содержится в плодах и овощах, остатки D-галактуроновой кислоты связаны альфа-1,4-гликозидными связями. В присутствии органических кислот спосбны к желеобразованию, применяются в пищевой промышленности для приготовления желе и мармелада. Некоторые пектиновые вещества оказывают противоязвенный эффект и являются активной составляющей ряда фармацевтических препаратов, например, производное подорожника «плантаглюцид»[2]. Мурами́н (лат. múrus — стенка) — полисахарид, опорно-механический материал клеточной стенки бактерий. По химическому строению представляет собой неразветвлённую цепь, построенную из чередующихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединённых бета-1,4-гликозидной связью. Мурамин по структурной организации (неразветвлённая цепь бета-1,4-полиглюкопиранозного скелета) и функциональной роли весьма близок к хитину и целлюлозе[2]. Декстра́ны — полисахариды бактериального происхождения — синтезируются в условиях промышленного производства микробиологическим путём (воздействием микроорганизмов Leuconostoc mesenteroides на раствор сахарозы) и используются в качестве заменителей плазмы крови (так называемые клинические «декстраны»: Полиглюкин и другие)[2]. В живых организмах углеводы выполняют следующие функции: 1. Структурная и опорная функции. Углеводы участвуют в построении различных опорных структур. Так целлюлоза является основным структурным компонентом клеточных стенок растений, хитин выполняет аналогичную функцию у грибов, а также обеспечивает жёсткость экзоскелета членистоногих[1]. 2. Защитная роль у растений. У некоторых растений есть защитные образования (шипы, колючки и др.), состоящие из клеточных стенок мёртвых клеток. 3. Пластическая функция. Углеводы входят в состав сложных молекул (например, пентозы (рибоза и дезоксирибоза) участвуют в построении АТФ, ДНК и РНК)[6]. 4. Энергетическая функция. Углеводы служат источником энергии: при окислении 1 грамма углеводов выделяются 4,1 ккал энергии и 0,4 г воды[6]. 5. Запасающая функция. Углеводы выступают в качестве запасных питательных веществ: гликоген у животных, крахмал и инулин — у растений[1]. 6. Осмотическая функция. Углеводы участвуют в регуляции осмотического давления в организме. Так, в крови содержится 100—110 мг/% глюкозы, от концентрации глюкозы зависит осмотическое давление крови. 7. Рецепторная функция. Олигосахариды входят в состав воспринимающей части многих клеточных рецепторов или молекул-лигандов. 7.Биологическое преобразование энергии. Фотосинтез, хемосинтез, дыхание Фотосинтез — это процесс образования органического вещества из углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилл у растений, бактериохлорофилл и бактериородопсин у бактерий). В современной физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества. Различают оксигенный и аноксигенный типы фотосинтеза. Оксигенный гораздо более широко распространён, его осуществляют растения, цианобактерии и прохлорофиты. В данной статье описан только он, аноксигенному фотосинтезу пурпурных и зелёных бактерий, а также геликобактерий посвящена отдельная статья. Выделяют три этапа фотосинтеза: фотофизический, фотохимический и химический. На первом этапе происходит поглощение квантов света пигментами, их переход в возбуждённое состояние и передача энергии к другим молекулам фотосистемы. На втором этапе происходит разделение зарядов в реакционном центре, перенос электронов по фотосинтетической электронотранспортной цепи, что заканчивается синтезом АТФ и НАДФН. Первые два этапа вместе называют светозависимой стадией фотосинтеза. Третий этап происходит уже без обязательного участия света и включает в себя биохимические реакции синтеза органических веществ с использованием энергии, накопленной на светозависимой стадии. Чаще всего в качестве таких реакций рассматривается цикл Кальвина и глюконеогенез, образование сахаров и крахмала из углекислого газа воздуха. Дыхание — основная форма диссимиляции у человека, животных, растений и многих микроорганизмов. При дыхании богатые энергией вещества, принадлежащие организму, полностью разлагаются до бедных энергией неорганических конечных продуктов (диоксида углерода и воды), используя для этого молекулярный кислород. Под внешним дыханием понимают газообмен между организмом и окружающей средой, включающий поглощение кислорода и выделение углекислого газа, а также транспорт этих газов внутри организма. Внутреннее (клеточное) дыхание включает биохимические процессы в цитоплазме клеток и митохондриях, приводящее к высвобождению энергии. У организмов, имеющих большие площади поверхности, контактирующие с внешней средой, дыхание может происходить за счёт диффузии газов непосредственно к клеткам (например, в листьях растений, у полостных животных). При небольшой относительной площади поверхности транспорт газов осуществляется за счёт циркуляции крови (у позвоночных и др.) либо в трахеях (у насекомых). Хемосинтез — способ автотрофного питания, при котором источником энергии для синтеза органических веществ из CO2 служат реакции окисления неорганических соединений. Подобный вариант получения энергии используется только бактериями. Явление хемосинтеза было открыто в 1887 году русским учёным С. Н. Виноградским. Необходимо отметить, что выделяющаяся в реакциях окисления неорганических соединений энергия не может быть непосредственно использована в процессах ассимиляции. Сначала эта энергия переводится в энергию макроэргических связей АТФ и только затем тратится на синтез органических соединений.
|
|||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-07; просмотров: 1836; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.222.98.91 (0.011 с.) |