
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Биофизика мышечного сокращенияСодержание книги
Похожие статьи вашей тематики
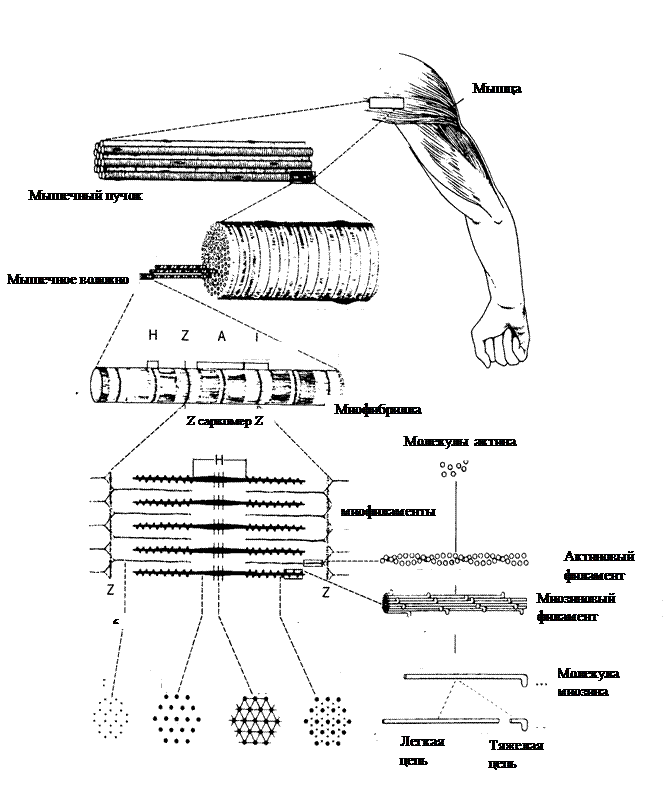
Поиск на нашем сайте БИОФИЗИКА МЫШЕЧНОГО СОКРАЩЕНИЯ Все разнообразные формы движений в природе, начиная с биения ресничек одноклеточных организмов и движения листьев растений и заканчивая сокращениями скелетных мышц, имеют некоторые общие черты. Все они связаны с превращением химической энергии, освобождающейся при гидролизе аденозинтрифосфата (АТФ), в механическую энергию. Кроме того, любые движения совершаются в клетках при участии специальных белковых молекул. Скелетные мышцы играют чрезвычайно важную роль в жизнедеятельности организма. Благодаря их сокращениям человек способен не только перемещаться в пространстве и поддерживать позу тела, но и выражать свои чувства посредством речи, письма и мимики. В настоящее время деятельность скелетных мышц изучена не только на клеточном, но и на молекулярном уровне, хотя многие принципы этой деятельности всё ещё остаются неизвестными. Для того чтобы понять биофизические механизмы мышечного сокращения, необходимо рассмотреть структуру скелетной мышцы. Структура скелетной мышцы Структура скелетной мышцы, включающая разные уровни её организации, представлена на рис. 1. Мышца состоит из отдельных пучков мышечной ткани, каждый из которых включает в себя большое число мышечных волокон - клеток, способных укорачиваться и совершать механическую работу. Скелетное мышечное волокно представляет собой многоядерную клетку, диаметр которой может составлять от 10 до 100 мкм, а длина её, как правило, равна длине мышцы в целом. Мышечные волокна окружены плазматической мембраной (сарколеммой), которая в состоянии покоя поляризована. Мембранный потенциал покоя скелетного мышечного волокна составляет около – 90мВ. Каждое мышечное волокно включает в себя от нескольких сот до двух тысяч более тонких вытянутых волоконец (миофибрилл) диаметром 1–2 мкм, участвующих непосредственно в мышечном сокращении. Миофибриллы погружены в протоплазму мышечного волокна (саркоплазму), в которой находятся обычные внутриклеточные органоиды. Основными ионами жидкости саркоплазмы яв -
Рис. 1. Структура скелетной мышцы
ляются катионы калия, анионы фосфата. Кроме того, в ней содержатся различные белки-ферменты. Отличительной чертой саркоплазмы мышечного волокна является наличие в ней множества митохондрий, синтезирующих большое количество АТФ. В саркоплазме имеются также две специализированные мембранные системы: Т-система и саркоплазматический ретикулум. Т-система представляет собой впячивания поверхностной мембраны в поперечном направлении внутрь мышечного волокна. Она контактирует с саркоплазматическим ретикулумом - системой вытянутых пузырьков, внутри которых содержатся в высокой концентрации ионы кальция. Миофибриллы состоят из меньших субъединиц – миофиламентов, которые представляют собой белковые молекулы, ответственные за сокращение мышцы. В мышце имеются два вида сократительных белков: актин и миозин. Каждая из миофибрилл содержит лежащие параллельно друг другу около 1500 миозиновых и 3000 актиновых филаментов. Миозиновые филаменты являются более толстыми (15нм), по сравнению с актиновыми (10нм). Миозиновые и актиновые филаменты частично перекрываются, в результате чего миофибриллы в микроскопе выглядят как совокупность светлых и тёмных дисков. Светлые диски содержат только актиновые филаменты и называются изотропными (I-диски). Темные диски, представленные перекрывающимися миозиновыми филаментами и расположенными между ними концами актиновых филаментов, называются анизотропными (А-диски). От поверхности миозиновых филаментов вдоль всей их длины (за исключением центральной части) отходят поперечные мостики. В покоящейся мышце концы миозиновых и актиновых филаментов обычно лишь слабо перекрываются на границе между А- и I- дисками. Эта зона перекрывания выглядит в световом микроскопе темнее центральной Н – зоны, в которой нет актиновых нитей. В середине саркомера (в центре Н-зоны) видна тонкая темная М –линия – сеть опорных белков, которые удерживают миозиновые филаменты в составе единого пучка. Концы актиновых волокон прикрепляются к, так называемой, Z-мембране, от которой они тянутся в обоих направлениях до мест перекрытия с миозиновыми филаментами. Мембрана Z проходитпоперёк каждой миофибриллы и соединяет все миофибриллы мышечного волокна друг с другом. Следовательно, мышечное волокно в целом имеет такие же светлые и тёмные диски, как и отдельные миофибриллы. Эти диски образуют повторяющиеся структурные элементы - саркомеры, которые придают скелетной мышце поперечную исчерченность, обнаруживаемую в миофибриллах с помощью светового микроскопа. Саркомер - часть миофибрилл (или мышечного волокна в целом), которая лежит между двумя соседними Z-мембранами. В покоящемся мышечном волокне длина саркомера составляет около 2,5мкм. Управление сократительной активностью скелетных мышц осуществляется аксонами двигательных нервных клеток - мотонейронов. Каждый из них иннервирует группу мышечных волокон, которые сокращаются как единое целое (нейромоторная единица). Число активных нейромоторных единиц в мышце может изменяться в зависимости от числа мотонейронов, активных в данный момент. От этого зависит сила и скорость сокращения мышцы. А) Б) В)
Рис. 7. Схематическое изображение типичных рычагов в теле человека: А) рычаг первого рода; Б) и В) рычаги второго рода. На рисунке
Рычаг – это твёрдое недеформируемое тело, имеющее точку опоры (вращения). В организме функцию рычагов выполняют кости скелета. На одно плечо рычага действует сила, развиваемая мышцей, а на другое – нагрузка, против которой работает мышца (чаще всего сила тяжести отдельных компонентов тела – головы, плеча, туловища и т.д.). Принято различать рычаги первого и второго рода. У рычага первого рода точка опоры расположена между линиями действующих сил. Примером рычага первого рода в теле служит шейно-затылочное сочленение и совокупность мышц, прикрепленных к основанию черепа спереди и сзади от него. Это сочленение можно рассматривать как точку опоры, вес головы спереди от сустава – как нагрузку (рис.7А). Уравновешивающая её сила мышцы, которая обеспечивает сохранение положения и движение черепа, располагается сзади от точки вращения. У рычага второго рода линии действующих сил находятся по одну сторону от точки опоры. Эти рычаги очень широко представлены в опорно-двигательном аппарате. Практически все элементы конечностей (кисть, предплечье, плечо, стопа, голень, бедро) являются рычагами второго рода. Примером такого рычага может служить система мышц, при помощи которых человек становится на носки (рис.7Б). В этой системе точкой опоры служат плюсневые кости стопы, нагрузкой – вес тела, приложенный к голеностопному суставу, а противодействующая ей сила направлена вверх и создаётся икроножной мышцей у места прикрепления Ахиллова сухожилия к пяточной кости. Еще одним примером рычага второго рода в теле служит локтевой сустав (рис.7В). Его можно рассматривать как точку опоры. При сгибании в суставе сила создаётся в результате сокращения двуглавой мышцы плеча. Эта сила приложена недалеко от точки опоры, а нагрузка создаётся весом предплечья и любого предмета, который человек держит в руке. Два рычага могут образовывать кинематическую пару, которая увеличивает объём их движения. Примером являются подвижные соединения костей (рычагов) - суставы. Более сложными системами из нескольких рычагов являются кинематические цепи (например, верхняя конечность, звеньями которой являются плечо, предплечье, кисть, фаланги пальцев). Объём движений в кинематических цепях значительно выше, чем в рычаге или кинематической паре. Строение организма человека характеризуется некоторыми особенностями, обеспечивающими определённые преимущества его опорно-двигательного аппарата. Примером может служить система прикрепления сухожилий сгибателей к фалангам пальцев. При сокращении этих мышц развиваемые ими силы прикладываются через сухожилия к фалангам пальцев. Система связок удерживает сухожилия в положении, приблизительно параллельном оси пальцев. При таком устройстве сокращение мышц приводит к сгибанию пальца, что позволяет захватывать предметы. Во многих случаях мышцы перекидываются не через один, а через два сустава. Это также создаёт определённые преимущества. Например, при беге нога одновременно сгибается в тазобедренном суставе и разгибается в коленном. При этом работу выполняет одна мышца, что позволяет получить экономию затрачиваемой энергии.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ: 1. Стейси Р., Уильямс Д., Уорден Р., Мак – Моррис Р. Основы биологической и медицинской физики. - М.: «Иностранная литература», 1959. - С.121. 2. Физиология человека: В 3-х томах. Т.1. /Под ред. Р. Шмидта и Г. Тевса. – М.: Мир, 1996. – С. 69-84. 3. Тиманюк В.А., Животова Е.Н. Биофизика. – К.: ИД «Профессионал», 2004. - С. 79-86. БИОФИЗИКА МЫШЕЧНОГО СОКРАЩЕНИЯ Все разнообразные формы движений в природе, начиная с биения ресничек одноклеточных организмов и движения листьев растений и заканчивая сокращениями скелетных мышц, имеют некоторые общие черты. Все они связаны с превращением химической энергии, освобождающейся при гидролизе аденозинтрифосфата (АТФ), в механическую энергию. Кроме того, любые движения совершаются в клетках при участии специальных белковых молекул. Скелетные мышцы играют чрезвычайно важную роль в жизнедеятельности организма. Благодаря их сокращениям человек способен не только перемещаться в пространстве и поддерживать позу тела, но и выражать свои чувства посредством речи, письма и мимики. В настоящее время деятельность скелетных мышц изучена не только на клеточном, но и на молекулярном уровне, хотя многие принципы этой деятельности всё ещё остаются неизвестными. Для того чтобы понять биофизические механизмы мышечного сокращения, необходимо рассмотреть структуру скелетной мышцы. Структура скелетной мышцы Структура скелетной мышцы, включающая разные уровни её организации, представлена на рис. 1. Мышца состоит из отдельных пучков мышечной ткани, каждый из которых включает в себя большое число мышечных волокон - клеток, способных укорачиваться и совершать механическую работу. Скелетное мышечное волокно представляет собой многоядерную клетку, диаметр которой может составлять от 10 до 100 мкм, а длина её, как правило, равна длине мышцы в целом. Мышечные волокна окружены плазматической мембраной (сарколеммой), которая в состоянии покоя поляризована. Мембранный потенциал покоя скелетного мышечного волокна составляет около – 90мВ. Каждое мышечное волокно включает в себя от нескольких сот до двух тысяч более тонких вытянутых волоконец (миофибрилл) диаметром 1–2 мкм, участвующих непосредственно в мышечном сокращении. Миофибриллы погружены в протоплазму мышечного волокна (саркоплазму), в которой находятся обычные внутриклеточные органоиды. Основными ионами жидкости саркоплазмы яв -
Рис. 1. Структура скелетной мышцы
ляются катионы калия, анионы фосфата. Кроме того, в ней содержатся различные белки-ферменты. Отличительной чертой саркоплазмы мышечного волокна является наличие в ней множества митохондрий, синтезирующих большое количество АТФ. В саркоплазме имеются также две специализированные мембранные системы: Т-система и саркоплазматический ретикулум. Т-система представляет собой впячивания поверхностной мембраны в поперечном направлении внутрь мышечного волокна. Она контактирует с саркоплазматическим ретикулумом - системой вытянутых пузырьков, внутри которых содержатся в высокой концентрации ионы кальция. Миофибриллы состоят из меньших субъединиц – миофиламентов, которые представляют собой белковые молекулы, ответственные за сокращение мышцы. В мышце имеются два вида сократительных белков: актин и миозин. Каждая из миофибрилл содержит лежащие параллельно друг другу около 1500 миозиновых и 3000 актиновых филаментов. Миозиновые филаменты являются более толстыми (15нм), по сравнению с актиновыми (10нм). Миозиновые и актиновые филаменты частично перекрываются, в результате чего миофибриллы в микроскопе выглядят как совокупность светлых и тёмных дисков. Светлые диски содержат только актиновые филаменты и называются изотропными (I-диски). Темные диски, представленные перекрывающимися миозиновыми филаментами и расположенными между ними концами актиновых филаментов, называются анизотропными (А-диски). От поверхности миозиновых филаментов вдоль всей их длины (за исключением центральной части) отходят поперечные мостики. В покоящейся мышце концы миозиновых и актиновых филаментов обычно лишь слабо перекрываются на границе между А- и I- дисками. Эта зона перекрывания выглядит в световом микроскопе темнее центральной Н – зоны, в которой нет актиновых нитей. В середине саркомера (в центре Н-зоны) видна тонкая темная М –линия – сеть опорных белков, которые удерживают миозиновые филаменты в составе единого пучка. Концы актиновых волокон прикрепляются к, так называемой, Z-мембране, от которой они тянутся в обоих направлениях до мест перекрытия с миозиновыми филаментами. Мембрана Z проходитпоперёк каждой миофибриллы и соединяет все миофибриллы мышечного волокна друг с другом. Следовательно, мышечное волокно в целом имеет такие же светлые и тёмные диски, как и отдельные миофибриллы. Эти диски образуют повторяющиеся структурные элементы - саркомеры, которые придают скелетной мышце поперечную исчерченность, обнаруживаемую в миофибриллах с помощью светового микроскопа. Саркомер - часть миофибрилл (или мышечного волокна в целом), которая лежит между двумя соседними Z-мембранами. В покоящемся мышечном волокне длина саркомера составляет около 2,5мкм. Управление сократительной активностью скелетных мышц осуществляется аксонами двигательных нервных клеток - мотонейронов. Каждый из них иннервирует группу мышечных волокон, которые сокращаются как единое целое (нейромоторная единица). Число активных нейромоторных единиц в мышце может изменяться в зависимости от числа мотонейронов, активных в данный момент. От этого зависит сила и скорость сокращения мышцы.
|
||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 695; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.214 (0.011 с.) |

 точка опоры рычага,
точка опоры рычага,  вес частей тела, стрелочкой обозначена линия действия силы, развиваемой мышцей
вес частей тела, стрелочкой обозначена линия действия силы, развиваемой мышцей


