
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
A 23 ионные теории возникновения пд (бернштейн и А. Ходжкин). Ионные каналы. Изменение проницаемости при генерации пд. Величина пд в разных тканях.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте Мембранная теория возбуждения, общепринятая в физиологии теория возбуждения мышечных и нервных клеток. Основа Мембранная теория возбуждения — представление о том, что при раздражении возбудимой клетки в её поверхностной мембране происходит молекулярная перестройка, которая приводит к изменению проницаемости мембраны и появлению трансмембранных ионных токов. Источником энергии для этих токов служит постоянно существующее неравномерное распределение основных неорганических ионов между цитоплазмой и внеклеточной средой: накопление в клетке ионов K+ и выведение из неё ионов Na+ и Cl- (рис. 1). Основные положения Мембранная теория возбуждения сформулированы немецким нейрофизиологом Ю. Бернштейном (1902) и развиты английскими учёными: П. Бойлом и Э. Конуэем (1941) и А. Ходжкином, Б. Кацем, А. Хаксли (1949). Бернштейн предположил, что поверхностная мембрана возбудимой клетки в покое обладает избирательной проницаемостью: ионы K+ проходят через неё гораздо легче, чем ионы Na+ и Cl-. Т. к. концентрация K+ в клетке выше, чем во внеклеточной среде, диффузия этих ионов через мембрану создаёт на ней разность потенциалов — т. н. потенциал покоя (ПП), причём внутренняя сторона мембраны оказывается заряженной отрицательно, а внешняя — положительно. (Зависимость ПП от ионов K+ подтверждается пропорциональным снижением его величины при увеличении содержания K+ во внеклеточной среде.) Чтобы объяснить, каким образом клетка поддерживает постоянный ионный состав и отрицательный внутренний потенциал, выводя ионы Na+ наружу, было выдвинуто предположение о возможности переноса ионов через мембрану не только под влиянием электрических сил и диффузии («пассивный» транспорт), но и посредством «активного» транспортного механизма — «натриевого насоса». В результате работы этого насоса, способного выталкивать Na+ против концентрационного и электрического градиентов, на каждый ион Na+, выбрасываемый через мембрану, клетка принимает один ион K+. При действии на клетку раздражения ионная проницаемость мембраны изменяется. Это обусловливается либо изменением электрического поля мембраны («электрическая» возбудимость), либо действием химических веществ на особые рецепторные структуры мембраны («химическая» возбудимость). По представлениям Бернштейна, при электрическом раздражении мембрана становится проницаемой для всех ионов, что приводит к кратковременному исчезновению ПП в возбуждённом участке — потенциалу действия (ПД). Последующие исследования (с применением микроэлектродной техники) явлений, возникающих при электрических раздражениях, показали, что ПД примерно в 1,5 раза превышает ПП. При этом происходит инверсия: возбуждённый участок мембраны приобретает разность потенциалов, противоположную по направлению той, какая существовала на ней в состоянии покоя (внутренняя сторона мембраны становится положительно заряженной по отношению к наружной). Однако при возбуждении происходит не общее (как думал Бернштейн), а избирательное увеличение ионной проницаемости мембраны — только для ионов Na+, которые проходят внутрь клетки, перенося через мембрану положительные заряды. Вследствие этого и возникает ПД. (Правильность такого объяснения подтверждается исчезновением ПД при устранении из внеклеточной среды Na+ при неизменном ПП, обнаружением потока ионов Na+ внутрь клетки при её возбуждении и т.д.). Наиболее точные данные об ионных токах через поверхностную мембрану при ПД получены методом т. н. фиксации напряжения на мембране. При этом одной парой электродов (один из них находится внутри клетки) измеряют разность потенциалов на мембране, а через др. пару пропускают ток от усилителя, поддерживающий эту разность на постоянном уровне, независимо от изменений в мембране. Т. о. было показано, что при возбуждении сначала возникает кратковременный ионный ток, направленный внутрь клетки, который затем сменяется ионным током, направленным наружу. Начальный, входящий ток обусловлен движением через мембрану Na+, выходящий — движением из клетки K+; в результате восстанавливается исходное состояние электрической поляризации клеточной мембраны. Кратковременность ионных токов, возникающих при ПД, связывают с наличием в мембране наряду с механизмом повышения («активации») ионной проницаемости также противоположного процесса — её «инактивации», обусловливающей развитие рефрактерности и аккомодации к электрическому раздражению. Появление в каком-либо участке возбудимой клетки ПД приводит к образованию на мембране «продольной» разности потенциалов и появлению электрических токов между невозбуждёнными и возбуждёнными участками — т. н. токов действия. Эти токи, в свою очередь, вызывают в невозбуждённых участках аналогичные изменения проницаемости; участок возбуждения начинает перемещаться по поверхности клетки (рис. 2). Описанные ионные процессы ведут (помимо появления распространяющегося импульса нервного) к накоплению в клетке некоторого количества Na+ и потере ею части K+. Эти изменения столь незначительны по сравнению с существующими между цитоплазмой и внеклеточной средой ионными градиентами, что клетка может генерировать огромное число импульсов без немедленного восстановления нарушенных ионных соотношений за счёт активного транспорта ионов, удаляющего из клетки избыток Na+ и насасывающего в неё недостающее количество K+. При химическом раздражении специфических изменения ионной проницаемости мембраны также приводят к развитию трансмембранных ионных токов. Такие изменения развиваются в межнейронных и нервно-мышечных синапсах и лежат в основе синаптической передачи с помощью медиаторов. Существо перестроек в мембране, обеспечивающих появление ионных токов, — наименее ясная часть Мембранная теория возбуждения Полагают, что перенос ионов через мембрану происходит либо по системе пор (входы в которые в состоянии покоя закрыты, возможно ионами Ca2+, и открываются под действием внешнего раздражения), либо при помощи особых молекул-переносчиков, которые связывают ион на одной стороне мембраны и освобождают его на другой A 24 Изменение МП при действии подпороговых раздражителей. Локальные ответы. Уровень критической деполяризации и порог деполяризации. Величина изменения мембранного потенциала клетки зависит от силы действующего на клетку раздражителя. При действии подпороговых раздражителей, реакция клетки ограничивается местными сдвигами мембранного потенциала которые получили название местных потенциалов (рис. 3.1 А, Б).
Рис. 3.1. Особенности возникновения и развития местного потенциала. А - Градуальность местного потенциала - чем сильнее раздражение, тем выше местный потенциал; местный потенциал не имеет определенного порога и возникает при любой силе раздражителя. Б - Продолжительность местного потенциала прямопропорциональна силе и длительности раздражения, местный потенциал не имеет латентного периода и возникает практически сразу после воздействия раздражителя. В - Местные потенциалы могут суммироваться. Так, если на фоне местного потенциала нанести новое подпороговое раздражение, то возникающий на второе раздражение ответ накладывается на первый и общий суммарный эффект от этого увеличивается. В возникновении и развитии местного потенциала можно выделить ряд специфических особенностей. Местный потенциал отличается отсутствием определенного порога и возникает он при действии подпороговых раздражителей малой силы. Другой особенностью местного потенциала является его градуальность, т.е. зависимость амплитуды ответа от силы раздражителя: чем сильнее раздражение, тем больше амплитуда местного потенциала. Продолжительность местного потенциала также непостоянна и зависит от силы и длительности раздражителя. Чем сильнее и длительнее раздражение, тем более продолжительное время сохраняется местный потенциал. Местный потенциал характеризуется отсутствием латентного периода и возникает практически сразу после воздействия раздражителя. Участок клеточной мембраны, на котором возникает местный потенциал, отличается повышенной возбудимостью. Кроме того, местный потенциал может суммироваться. Так, если на фоне местного потенциала нанести новое подпороговое раздражение, возникший на второе раздражение ответ накладывается на первый, и общий суммарный эффект от этого увеличивается (рис.3.1 В). Исследование механизма возникновения местных потенциалов показало, что они могут быть результатом как пассивных, так и активных процессов в клеточной мембране. Изменение мембранно-го потенциала при протекании через мембрану электрического тока от постоянного источника не зависит от молекулярных процессов в самой мембране. Такое изменение потенциала, не связанное с включением ионных каналов, является результатом пассивных процессов (рис. 3.2).
Рис. 3.2. Возникновение местного потенциала при прохождении электрического тока, через участок нерва. Ток идет от анода к катоду (оба электрода - вне нерва) частично через пленку жидкости на поверхности нерва, а частично через оболочку нерва и в продольном направлении внутри волокна. Кривая внизу показывает вызываемое током изменения мембранного потенциала нервного волокна (по В. Катц). У анода положительные заряды, поступающие к наружной поверхности мембраны, увеличивают ее внешний положительный заряд. Внутренний отрицательный заряд на этом участке также увеличивается вследствие перемещения положительных ионов к катоду. В результате этого мембрана у анода гиперполяризуется. Противоположный сдвиг - деполяризация происходит у катода. Такие изменения мембранного потенциала, не связанные с изменениями ионной проницаемости мембраны, принято называть пассивными или электротоническими. В том случае, когда сила действующего допорогового стимула составляет более 50-75% от пороговой величины в изменении мембранного потенциала играют роль и активные процессы (рис.3.3). При этом изменяется проницаемость мембраны для натрия за счет активации натриевых каналов; такие изменения являются активными и называются локальным ответом.
Амплитуда локального ответа увеличивается по мере приближения силы раздражителя к порогу, а при его достижении локальный ответ перерастает в потенциал действия.
|
||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 1408; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.214 (0.008 с.) |

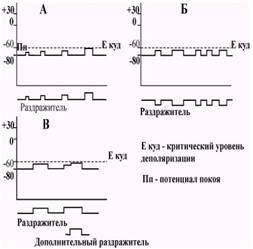
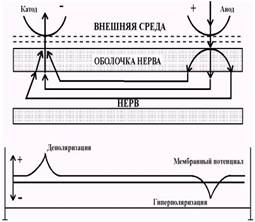
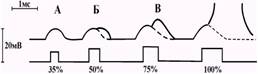 Рис. 3.3. Вклад пассивных и активных процессов в развитие местного потенциала, при действии допороговых раздражителей разной силы.
Рис. 3.3. Вклад пассивных и активных процессов в развитие местного потенциала, при действии допороговых раздражителей разной силы. 


