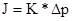Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Розділ 3. Основи мембранних процесівСодержание книги
Поиск на нашем сайте
Зворотний осмос та ультрафільтрація мають подібності (матеріал мембран) та відмінності (діаметр пор в мембранах, тому різний механізм процесів). Осмос Осмос (от греч. ōsmós — толчок, давление), диффузия вещества, обычно растворителя, через полупроницаемую мембрану, разделяющую раствор и чистый растворитель или два раствора различной концентрации. Полупроницаемая мембрана — перегородка, пропускающая малые молекулы растворителя, но непроницаемая для более крупных молекул растворённого вещества. Выравнивание концентраций по обе стороны такой мембраны возможно лишь при односторонней диффузии растворителя. Поэтому всегда идёт от чистого растворителя к раствору или от разбавленного раствора к концентрированному. О., направленный внутрь ограниченного объёма жидкости, называется эндосмосом, наружу — экзосмосом. Перенос растворителя через мембрану обусловлен осмотическим давлением. Оно равно избыточному внешнему давлению, которое следует приложить со стороны раствора, чтобы прекратить О., т. е. создать условия осмотического равновесия. Превышение избыточного давления над осмотическим может привести к обращению О. — обратной диффузии растворителя (см. Ультрафильтрация). В случаях, когда мембрана проницаема не только для растворителя, но и для некоторых растворённых веществ, диффузия последних из раствора в растворитель позволяет осуществить диализ, применяемый как способ очистки полимеров и коллоидных систем от низкомолекулярных примесей, например электролитов. Впервые О. наблюдал А. Нолле в 1748, однако исследование этого явления было начато спустя столетие. О. имеет важнейшее значение в биологических процессах (см. Осморегуляция), его широко используют в лабораторной технике: при определении молярных характеристик полимеров, концентрировании растворов, исследовании разнообразных биологических структур. Осмотические явления иногда используются в промышленности, например при получении некоторых полимерных материалов, очистке высоко-минерализованной воды методом "обратного" О. жидкостей. Осмотическое давление Осмометр (схема) Осмотическое давление, диффузное давление, термодинамический параметр, характеризующий стремление раствора к понижению концентрации при соприкосновении с чистым растворителем вследствие встречной диффузии молекул растворённого вещества и растворителя. Если раствор отделен от чистого растворителя полупроницаемой мембраной, то возможна лишь односторонняя диффузия — осмотическое всасывание растворителя через мембрану в раствор. В этом случае О. д. становится доступной для прямого измерения величиной, равной избыточному давлению, приложенному со стороны раствора при осмотическом равновесии (см. Осмос). О. д. обусловлено понижением химического потенциала растворителя в присутствии растворённого вещества. Тенденция системы выравнивать химические потенциалы во всех частях своего объёма и перейти в состояние с более низким уровнем свободной энергии вызывает осмотическое (диффузионный) перенос вещества. О. д. в идеальных и предельно разбавленных растворах не зависит от природы растворителя и растворённых веществ; при постоянной температуре оно определяется только числом "кинетических элементов" — ионов, молекул, ассоциатов или коллоидных частиц — в единице объёма раствора. Первые измерения О. д. произвёл В. Пфеффер (1877), исследуя водные растворы тростникового сахара. Его данные позволили Я. X. Вант-Гоффу установить (1887) зависимость О. д. от концентрации растворённого вещества, совпадающую по форме с Бойля — Мариотта законом для идеальных газов. Оказалось, что О. д. (p) численно равно давлению, которое оказало бы растворённое вещество, если бы оно при данной температуре находилось в состоянии идеального газа и занимало объём, равный объёму раствора. Для весьма разбавленных растворов недиссоциирующих веществ найденная закономерность с достаточной точностью описывается уравнением: pV = nRT, где n — число молей растворённого вещества в объёме раствора V; R — универсальная газовая постоянная; Т — абсолютная температура. В случае диссоциации вещества в растворе на ионы в правую часть уравнения вводится множитель i > 1, коэффициент Вант-Гоффа; при ассоциации растворённого вещества i < 1. О. д. реального раствора (p') всегда выше, чем идеального (p''), причём отношение p'/ p'' = g, называемое осмотическим коэффициентом, увеличивается с ростом концентрации. Растворы с одинаковым О. д. называется изотоническими или изоосмотическими. Так, различные кровезаменители и физиологические растворы изотоничны относительно внутренних жидкостей организма. Если один раствор в сравнении с другим имеет более высокое О. д., его называют гипертоническим, а имеющий более низкое О. д. — гипотоническим. О. д. измеряют с помощью специальных приборов — осмометров. Различают статические и динамические методы измерения. Первый метод основан на определении избыточного гидростатического давления по высоте столба жидкости Н в трубке осмометров (рис.) после установления осмотического равновесия при равенстве внешних давлений p A и р Б в камерах А и Б. Второй метод сводится к измерению скоростей v всасывания и выдавливания растворителя из осмотической ячейки при различных значениях избыточного давления D p = p A — р Б с последующей интерполяцией полученных данных к n = 0 при D p = p. Многие осмометры позволяют использовать оба метода. Одна из главных трудностей в измерении О. д. — правильный подбор полупроницаемых мембран. Обычно применяют плёнки из целлофана, природных и синтетических полимеров, пористые керамические и стеклянные перегородки. Учение о методах и технике измерения О. д. называются осмометрией. Основное приложение осмометрии — определение молекулярной массы (М) полимеров. Значения М вычисляют из соотношения О. д. может достигать значительных величин. Например, 4%-ный раствор сахара при комнатной температуре имеет О. д. около 0,3 Мн/м 2, а 53%-ный — около 10 Мн/м 2; О. д. морской воды около 0,27 Мн/м 2. Л. А. Шиц. О. д. в клетках животных, растений, микроорганизмов и в биологических жидкостях зависит от концентрации веществ, растворённых в их жидких средах. Солевой состав биологических жидкостей и клеток, характерный для организмов каждого вида, поддерживается избирательной проницаемостью биологических мембран для разных солей и активным транспортом ионов. Относительное постоянство О. д. обеспечивается водно-солевым обменом, т. е. всасыванием, распределением, потреблением и выделением воды и солей (см. Выделение, Выделительная система, Осморегуляция). У т. н. гиперосмотических организмов внутреннего О. д. больше внешнего, у гипоосмотических — меньше внешнего; у изоосмотических (пойкилоосмотических) внутреннее О. д. равно внешнему. В первом случае ноны активно поглощаются организмом и задерживаются в нём, а вода поступает через биологич. мембраны пассивно, в соответствии с осмотическим градиентом. Гиперосмотическая регуляция свойственна пресноводным организмам, мор. хрящевым рыбам (акулы, скаты) и всем растениям. У организмов с гипоосмотической регуляцией имеются приспособления для активного выделения солей. У костистых рыб преобладающие в океанических водах ионы Na+ и Cl— выделяются через жабры, у морских пресмыкающихся (змеи и черепахи) и у птиц — через особые солевые железы, расположенные в области головы. Ионы Mg2+, Диапазон средних величин О. д. в клетках организмов, не способных поддерживать осмотический гомеостаз, довольно широк и зависит от вида и возраста организма, типа клеток и О. д. окружающей среды. В оптимальных условиях О. д. клеточного сока наземных органов болотных растений колеблется от 2 до 16 ат, у степных — от 8 до 40 ат. В разных клетках растения О. д. может резко различаться (так, у мангровых О. д. клеточного сока около 60 ат, а О. д. в сосудах ксилемы не превышает 1—2 ат). У гомойосмотических организмов, т. е. способных поддерживать относительное постоянство О. д., средней величины и диапазон колебаний О. д. различны (дождевой червь — 3,6—4,8 ат, пресноводные рыбы — 6,0—6,6, океанические костистые рыбы — 7,8—8,5, акуловые — 22,3—23,2, млекопитающие — 6,6—8,0 ат). У млекопитающих О. д. большинства биологических жидкостей равно О. д. крови (исключение составляют жидкости, выделяемые некоторыми железами, — слюна, пот, моча и др.). О. д., создаваемое в клетках животных высокомолекулярными соединениями (белки, полисахариды и др.), незначительно, но играет важную роль в обмене веществ (см. Онкотическое давление). Ю. В. Наточин, В. В. Кабанов. Лит.: Мелвин-Хьюз Э. А., Физическая химия, пер. с англ., кн. 1—2, М., 1962; Курс физической химии, под ред. Я. И. Герасимова, т. 1—2, М. — Л., 1963—1966; Пасынский А. Г., Коллоидная химия, 3 изд., М., 1968: Проссер Л., Браун Ф., Сравнительная физиология животных, пер. с англ., М., 1967; Гриффин Д., Новик Эл., Живой организм, пер. с англ., 1973; Нобел П., Физиология растительной клетки (физико-химический подход), пер. с англ., М., 1973.
Теорія ультрафільтрації. Ультрафільтрація – мембранний процес для відділення високомолекулярних речовин від низькомолекулярних. Мембрани пористі асиметричні, розмір пор мембрани 1…100 нм. Рушійна сила – тиск (1…5 бар). Принцип розділення – ситовий механізм. Розмір видаляємих часток 0,005-0,05 мкм. Потік через ультрафільтрацій мембрану пропорційний прикладеному тиску:
К – константа проникності, яка включає всі види структурних факторів. К=0,1…10 м3/м2*сут*бар (щільні…більш відкриті мембрани). рителем вследствие встречной диффузии молекул растворённого вещества и растворителя. Если раствор отделен от чистого растворителя полупроницаемой мембраной, то возможна лишь односторонняя диффузия — осмотическое всасывание растворителя через мембрану в раствор. В этом случае О. д. становится доступной для прямого измерения величиной, равной избыточному давлению, приложенному со стороны раствора при осмотическом равновесии (см. Осмос). О. д. обусловлено понижением химического потенциала растворителя в присутствии растворённого вещества. Тенденция системы выравнивать химические потенциалы во всех частях своего объёма и перейти в состояние с более низким уровнем свободной энергии вызывает осмотическое (диффузионный) перенос вещества. О. д. в идеальных и предельно разбавленных растворах не зависит от природы растворителя и растворённых веществ; при постоянной температуре оно определяется только числом "кинетических элементов" — ионов, молекул, ассоциатов или коллоидных частиц — в единице объёма раствора. Первые измерения О. д. произвёл В. Пфеффер (1877), исследуя водные растворы тростникового сахара. Его данные позволили Я. X. Вант-Гоффу установить (1887) зависимость О. д. от концентрации растворённого вещества, совпадающую по форме с Бойля — Мариотта законом для идеальных газов. Оказалось, что О. д. (p) численно равно давлению, которое оказало бы растворённое вещество, если бы оно при данной температуре находилось в состоянии идеального газа и занимало объём, равный объёму раствора. Для весьма разбавленных растворов недиссоциирующих веществ найденная закономерность с достаточной точностью описывается уравнением: pV = nRT, где n — число молей растворённого вещества в объёме раствора V; R — универсальная газовая постоянная; Т — абсолютная температура. В случае диссоциации вещества в растворе на ионы в правую часть уравнения вводится множитель i > 1, коэффициент Вант-Гоффа; при ассоциации растворённого вещества i < 1. О. д. реального раствора (p') всегда выше, чем идеального (p''), причём отношение p'/ p'' = g, называемое осмотическим коэффициентом, увеличивается с ростом концентрации. Растворы с одинаковым О. д. называется изотоническими или изоосмотическими. Так, различные кровезаменители и физиологические растворы изотоничны относительно внутренних жидкостей организма. Если один раствор в сравнении с другим имеет более высокое О. д., его называют гипертоническим, а имеющий более низкое О. д. — гипотоническим. О. д. измеряют с помощью специальных приборов — осмометров. Различают статические и динамические методы измерения. Первый метод основан на определении избыточного гидростатического давления по высоте столба жидкости Н в трубке осмометров (рис.) после установления осмотического равновесия при равенстве внешних давлений p A и р Б в камерах А и Б. Второй метод сводится к измерению скоростей v всасывания и выдавливания растворителя из осмотической ячейки при различных значениях избыточного давления D p = p A — р Б с последующей интерполяцией полученных данных к n = 0 при D p = p. Многие осмометры позволяют использовать оба метода. Одна из главных трудностей в измерении О. д. — правильный подбор полупроницаемых мембран. Обычно применяют плёнки из целлофана, природных и синтетических полимеров, пористые керамические и стеклянные перегородки. Учение о методах и технике измерения О. д. называются осмометрией. Основное приложение осмометрии — определение молекулярной массы (М) полимеров. Значения М вычисляют из соотношения О. д. может достигать значительных величин. Например, 4%-ный раствор сахара при комнатной температуре имеет О. д. около 0,3 Мн/м 2, а 53%-ный — около 10 Мн/м 2; О. д. морской воды около 0,27 Мн/м 2. Л. А. Шиц. О. д. в клетках животных, растений, микроорганизмов и в биологических жидкостях зависит от концентрации веществ, растворённых в их жидких средах. Солевой состав биологических жидкостей и клеток, характерный для организмов каждого вида, поддерживается избирательной проницаемостью биологических мембран для разных солей и активным транспортом ионов. Относительное постоянство О. д. обеспечивается водно-солевым обменом, т. е. всасыванием, распределением, потреблением и выделением воды и солей (см. Выделение, Выделительная система, Осморегуляция). У т. н. гиперосмотических организмов внутреннего О. д. больше внешнего, у гипоосмотических — меньше внешнего; у изоосмотических (пойкилоосмотических) внутреннее О. д. равно внешнему. В первом случае ноны активно поглощаются организмом и задерживаются в нём, а вода поступает через биологич. мембраны пассивно, в соответствии с осмотическим градиентом. Гиперосмотическая регуляция свойственна пресноводным организмам, мор. хрящевым рыбам (акулы, скаты) и всем растениям. У организмов с гипоосмотической регуляцией имеются приспособления для активного выделения солей. У костистых рыб преобладающие в океанических водах ионы Na+ и Cl— выделяются через жабры, у морских пресмыкающихся (змеи и черепахи) и у птиц — через особые солевые железы, расположенные в области головы. Ионы Mg2+, Диапазон средних величин О. д. в клетках организмов, не способных поддерживать осмотический гомеостаз, довольно широк и зависит от вида и возраста организма, типа клеток и О. д. окружающей среды. В оптимальных условиях О. д. клеточного сока наземных органов болотных растений колеблется от 2 до 16 ат, у степных — от 8 до 40 ат. В разных клетках растения О. д. может резко различаться (так, у мангровых О. д. клеточного сока около 60 ат, а О. д. в сосудах ксилемы не превышает 1—2 ат). У гомойосмотических организмов, т. е. способных поддерживать относительное постоянство О. д., средней величины и диапазон колебаний О. д. различны (дождевой червь — 3,6—4,8 ат, пресноводные рыбы — 6,0—6,6, океанические костистые рыбы — 7,8—8,5, акуловые — 22,3—23,2, млекопитающие — 6,6—8,0 ат). У млекопитающих О. д. большинства биологических жидкостей равно О. д. крови (исключение составляют жидкости, выделяемые некоторыми железами, — слюна, пот, моча и др.). О. д., создаваемое в клетках животных высокомолекулярными соединениями (белки, полисахариды и др.), незначительно, но играет важную роль в обмене веществ (см. Онкотическое давление). Ю. В. Наточин, В. В. Кабанов. Лит.: Мелвин-Хьюз Э. А., Физическая химия, пер. с англ., кн. 1—2, М., 1962; Курс физической химии, под ред. Я. И. Герасимова, т. 1—2, М. — Л., 1963—1966; Пасынский А. Г., Коллоидная химия, 3 изд., М., 1968: Проссер Л., Браун Ф., Сравнительная физиология животных, пер. с англ., М., 1967; Гриффин Д., Новик Эл., Живой организм, пер. с англ., 1973; Нобел П., Физиология растительной клетки (физико-химический подход), пер. с англ., М., 1973.
Теорія ультрафільтрації. Ультрафільтрація – мембранний процес для відділення високомолекулярних речовин від низькомолекулярних. Мембрани пористі асиметричні, розмір пор мембрани 1…100 нм. Рушійна сила – тиск (1…5 бар). Принцип розділення – ситовий механізм. Розмір видаляємих часток 0,005-0,05 мкм. Потік через ультрафільтрацій мембрану пропорційний прикладеному тиску:
К – константа проникності, яка включає всі види структурних факторів. К=0,1…10 м3/м2*сут*бар (щільні…більш відкриті мембрани).
[1] Апірогенна (а - не та прос – вогонь) вода – вода приготовлена на ХВО, яка не викликає у людини температури, необхідна для виготовлення ліків. [2] Взважений шар – киплячий шар, в якому частки твердого матеріалу інтенсивно пересуваються відносно одна одної під впливом потоку газу чи рідини, яка крізь нього проходить та мають дуже щільний контакт з цим потоком. Шар нагадує киплячу рідину та має деякі її властивості, підкоряється законам гідростатики. [3] Радіус іонів, Å: Н 1,54; О 0,096+ (1,46 [заряд 2-]); Ca 0,94; Na 0,98; Mg 0,65 [4] Спочатку припускали, що частка іоніту оточена плівкою рідини, яку вона міцно утримує та залишається нерухомою при будь-якому перемішування розчину. Зараз наявність такої нерухомої плівки відкидається, а взамін вводиться уявлення про зону рідини певної товщини навколо частки іоніту, в якій конвекція не впливає на розподілення концентрації іонів і остання обумовлена лише дифузійними процесами. [5] Броунівський рух – рух взважених в рідині часток, який викликаний безпорядочними ударами молекул навколишнього середовища Дифузія в розчинах – природний процес, який приводить до рівномірному розподіленню розчиненої речовини по об'єму розчину [6] Вуглеводні – з’єднання, молекули яких складаються з вуглецю С та водню Н. [7] Містять карбонову групу СООН – оцтова CH3COOH, мурашина HCOOH, бензойна, саліцилова. [8] Аміни – похідні аміаку –NH2CH3, NH(CH3)2 [9] Розподілення Гауса – нормальне розподілення – розподілення ймовірностей випадкової величини – виникає, якщо випадкова величина дорівнює сумі великого числа незалежних випадкових величин; ймовірність появи величини в певному інтервалі. [10] Бензол – добавка до палива для підвищення октанового числа [11] Ангстрем 1Å =10-10 м. Діаметр атому приблизно дорівнює ангстрему [12] Див. п.4.5 конспекту [13] Аміноз’єднання або аміни - органічні похідні аміаку. Аміни м.б. первинними R-NH2, вторинними RR'NH, третинними RR'R''N в залежності від числа атомів водню, які заміщені на радикали R, R', R''. Н-д, первинний амін – етіламін C2H5NH2, вторинний – діметиламін (CH3)2NH, третинний – триетіламін (C2H5)3N. Аміни виявляють основні властивості, в водному розчині гідратуються та дисоціюють як слабкі основи. Четвертинні амонієві групи –СH2Cl [14] Аміногрупи – первинні NH2 та вторинні NH – характерні групування амінів, мають пірамідальну будову. Проявляють відносно сильні основні властивості. [15] Насичені гетероциклічні з’єднання з групою NH у кільці. Сильні основи.
|
||||
|
|
Последнее изменение этой страницы: 2017-02-05; просмотров: 186; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.139.108.99 (0.01 с.) |

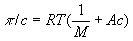 , где с — концентрация полимера по массе; А — коэффициент, зависящий от строения макромолекулы.
, где с — концентрация полимера по массе; А — коэффициент, зависящий от строения макромолекулы. ,
, 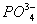 у этих организмов выделяются через почки. О. д. у гипер- и гипоосмотических организмов может создаваться как за счёт ионов, преобладающих во внешней среде, так и продуктов обмена. Например, у акуловых рыб и скатов О. д. на 60% создаётся за счёт мочевины и триметиламмония; в плазме крови млекопитающих — главным образом за счёт ионов Na+ и Cl—; в личинках насекомых — за счёт разнообразных низкомолекулярных метаболитов. У морских одноклеточных, иглокожих, головоногих моллюсков, миксин и др. изоосмотических организмов, у которых О. д. определяется О. д. внешней среды и равно ему, механизмы осморегуляции отсутствуют (исключая клеточные).
у этих организмов выделяются через почки. О. д. у гипер- и гипоосмотических организмов может создаваться как за счёт ионов, преобладающих во внешней среде, так и продуктов обмена. Например, у акуловых рыб и скатов О. д. на 60% создаётся за счёт мочевины и триметиламмония; в плазме крови млекопитающих — главным образом за счёт ионов Na+ и Cl—; в личинках насекомых — за счёт разнообразных низкомолекулярных метаболитов. У морских одноклеточных, иглокожих, головоногих моллюсков, миксин и др. изоосмотических организмов, у которых О. д. определяется О. д. внешней среды и равно ему, механизмы осморегуляции отсутствуют (исключая клеточные).