Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Передача энергии в экосистемеСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Самостоятельная работа № 1 Передача энергии в экосистеме Согласно закону максимизации энергии (Г. и Э. Одумов, 1972) выживает та экосистема, которая обеспечивает максимальное поступление и наиболее эффективное использование энергии. Поток энергии в экосистемах осуществляется через сложный механизм, включающий процессы ассимиляции, накопления, транспорт и высвобождение энергии. Важнейший этап в этом механизме - ассимиляция энергии, которая представляет собой превращение кинетической энергии фотонов в биологически доступную потенциальную энергию химических связей в органических соединениях. По способности к ассимиляции энергии организмы делятся на два типа: - автотрофы (продуценты) - организмы, самостоятельно синтезирующие органические вещества, поглощая минеральные вещества и энергию, поступающую из вне; - гетеротрофы (консументы и редуценты) – организмы, получающие необходимые органические вещества и энергию с пищей, представляющую собой живую или мертвую биомассу. К автотрофам относятся зеленые растения и некоторые виды бактерий. Гетеротрофами являются все животные, грибы, подавляющее большинство бактерий, некоторые водоросли и бесхлорофилльные высшие растения. Трофические цепи и сети Транспорт энергии в экосистемах осуществляется через пищевые цепи. Пищевая цепь – это перенос энергии в форме пищи от ее источника (автотрофа) через ряд организмов, происходящий путем поедания одних организмов другими. Обычно различают два типа пищевых цепей: - пастбищная цепь, начинающаяся живыми растениями (продуцентами), живой тканью которых питаются растительноядные животные (фитофаги); - детритная цепь, начинающаяся мертвым органическим веществом (детритом), потребляемым мертвоядными организмами (детритофагами).
Рис. 2 - Y-образная модель потока энергии, показывающая связь между пастбищной и детритной пищевыми цепями (Ю. Одум, 1986) Различают два типа пищевых цепей (рис. 2). Цепи выедания (или пастбищные) — пищевые цепи, начинающиеся с живых фотосинтезирующих организмов. Например, фитопланктон —» зоопланктон—> рыбы микрофаги —» рыбы макрофаги —> птицы ихтиофаги. Цепи разложения (или детритные) — пищевые цепи, начинающиеся с отмерших остатков растений, трупов и экскрементов животных. Например, детрит —» детритофаги —> хищники микрофаги —» хищники макрофаги. Таким образом, поток энергии, проходящий через экосистему, разбивается как бы на два основных направления. Энергия к консументам поступает через живые ткани растений или через запасы мертвого органического вещества. Цепи выедания преобладают в водных экосистемах, цепи разложения — в экосистемах суши. Передача энергии по пищевой цепи подчиняется второму закону термодинамики, согласно которому процессы, связанные с превращениями энергии, могут происходить самопроизвольно только при условии, что часть этой энергии рассеивается в форме тепла. В связи с этим, в каждом звене пищевой цепи теряется значительная доля (80-95%) энергии, заключенной в доступной пище. В сообществах пищевые цепи сложным образом переплетаются и образуют пищевые сети. В состав пищи каждого вида входит обычно не один, а несколько видов, каждый из которых в свою очередь может служить пищей нескольким видам. С одной стороны, каждый трофический уровень представлен многими популяциями разных видов, с другой стороны, многие популяции принадлежат сразу к нескольким трофическим уровням. В результате благодаря сложности пищевых связей выпадение какого-то одного вида часто не нарушает равновесия в экосистеме. Экологические пирамиды Закономерности транспорта энергии в пищевых цепях удобно изображать графически с помощью диаграмм, которые называются «экологические пирамиды». Они представляют собой соотношение численности организмов, их биомасс, эквивалентных этим биомассам энергий или продукции организмов на различных трофических уровнях пищевой цепи. Существует три основных типа пирамид -пирамида чисел - показывает численность отдельных организмов; -пирамида биомассы - характеризует общий сухой вес, калорийность или другую меру общей численности живого вещества; -пирамида энергии отвечает величине потока энергии или продуктивности на последовательных трофических уровнях.
Рис 3 - Пирамиды чисел (а), биомасс (б) и энергии (в), представляющие упрощенную экосистему: люцерна — телята — мальчик 12 лет (по Ю. Одуму, 1959) Пирамиды чисел и биомассы могут быть обратными, т.е. основа может быть меньшей, чем один или несколько верхних уровней. Так бывает когда средние размеры продуцентов меньше чем размеры консументов. Пирамида энергии всегда сужается к верху при условии, что будут учтены все источники питания в системе. Самостоятельная работа № 2 Закон толерантности Всю сложность взаимоотношения экологических факторов отражает закон толерантности В. Шелфорда: отсутствие или невозможность процветания определяется недостатком (в качественном или количественном смысле) или, наоборот, избытком любого из ряда факторов, уровень которых может оказаться близким к пределам переносимого данным организмом. Эти два предела называют пределами толерантности. Например, организм способен существовать при температуре от минус 5° С до плюс 25°С. Это и будет диапазоном толерантности организма по отношению к температуре. Факторы среды имеют количественное выражение (рис.). По отношению к каждому фактору можно выделить зону оптимума (зону нормальной жизнедеятельности), зону пессимума (зону угнетения) и пределы выносливости организма.
Рис - Зависимость действия экологического фактора от его количества
Рис. - Экологическая валентность (пластичность) видов: 1 - эврибионтные; 2 - стенобионтные Оптимум — такое количество экологического фактора, при котором интенсивность жизнедеятельности организмов максимальна. В зоне пессимума жизнедеятельность организмов угнетена. За пределами выносливости существование организма невозможно. Различают нижний и верхний предел выносливости. Способность живых организмов переносить количественные колебания действия экологического фактора в той или иной степени называется экологической валентностью (толерантностью, устойчивостью, пластичностью). Значения экологического фактора между верхним и нижним пределами выносливости называется зоной толерантности. Виды с широкой зоной толерантности называются эврибионтными, с узкой - стенобионтными. Организмы, переносящие значительные колебания температуры, называются эвритермные, а приспособленные к узкому интервалу температур - стенотермные. Таким же образом по отношению к давлению различают эври- и стенобатные организмы, по отношению к степени засоления среды - эври- и стеногалинные, и т.д. Экологические валентности отдельных индивидуумов не совпадают. Поэтому экологическая валентность вида шире экологической валентности каждой отдельной особи.
Рис - Экологическая валентность (пластичность) видов (по Ю. Одуму, 1975) Экологические валентности вида к разным экологическим факторам могут существенно отличаться. Набор экологических валентностей по отношению к разным факторам среды составляет экологический спектр вида. Лимитирующие факторы Лимитирующими (ограничивающими) экологическими факторами следует называть такие факторы, которые ограничивают развитие организмов из-за недостатка или их избытка по сравнению с потребностью (оптимальным содержанием). Такой фактор будет ограничивать распространение вида даже в том случае, если все остальные факторы будут благоприятными. Лимитирующие факторы определяют географический ареал вида. Знание человеком лимитирующих факторов для того или иного вида организмов позволяет, изменяя условия среды обитания, либо подавлять, либо стимулировать его развитие. В середине XIX в. Ю. Либихом был установлен закон минимума: урожай зависит от фактора, находящегося в минимуме. Например, если фосфор содержится в почве лишь в минимальных количествах, то это снижает урожай. Но оказалось, что если это же вещество находится в избытке, это также снижает урожай. Более того, факторы могут действовать изолированно или совокупно - ведь урожай зависит и от влажности, и от других факторов жизни растений. Тем не менее, факторы не могут заменить друг друга, что и нашло отражение в законе независимости факторов В.Р. Вильямса: условия жизни равнозначны, ни один из факторов жизни не может быть заменен другим. Например, нельзя заменить действие влажности действием углекислого газа или солнечного света и т. п. Можно выделить основные закономерности действия экологических факторов: закон относительности действия экологического фактора - направление и интенсивность действия экологического фактора зависят от того, в каких количествах он берется и в сочетании с какими другими факторами действует. Не бывает абсолютно полезных или вредных экологических факторов: все дело в количестве. Например, если температура окружающей среды слишком низкая или слишком высокая, т.е. выходит за пределы выносливости живых организмов, это для них плохо. Благоприятными являются только оптимальные значения. При этом экологические факторы нельзя рассматривать в отрыве друг от друга. Например, если организм испытывает дефицит воды, то ему труднее переносить высокую температуру; закон относительной заменяемости и абсолютной незаменимости экологических факторов - абсолютное отсутствие какого-либо из обязательных условий жизни заменить другими экологическими факторами невозможно, но недостаток или избыток одних экологических факторов может быть возмещен действием других экологических факторов. Например, полное (абсолютное) отсутствие воды нельзя компенсировать другими экологическими факторами. Однако если другие экологические факторы находятся в оптимуме, то перенести недостаток воды легче, чем когда и другие факторы находятся в недостатке или избытке.
Самостоятельная работа № 3 Структура биоценоза Пространственная структура - распределение организмов разных видов в пространстве (по вертикали и по горизонтали). Пространственная структура образуется прежде всего растительной частью биоценоза. Различают ярусность (вертикальная структура биоценоза) и мозаичность (структура биоценоза по горизонтали). Экологическая структура - соотношение организмов разных экологических групп. Биоценозы со сходной экологической структурой могут иметь разный видовой состав. Это связано с тем, что одни и те же экологические ниши могут быть заняты сходными по экологии, но далеко не родственными видами. Такие виды называются замещающими или викарирующими. Важными характеристиками структуры биоценоза являются консорция, синузия и парцелла. Консорция - структурная единица биоценоза, объединяющая автотрофные и гетеротрофные организмы на основе пространственных (топических) и пищевых (трофических) связей вокруг центрального члена (ядра). Например, отдельно стоящее дерево или группа деревьев (растение-эдификатор) и связанные с ним организмы. Биоценоз - это система связанных между собой консорций.
Рис. Консорция, парцелла и синузия (Н.Ф. Реймерс, 1990) Синузия - структурная часть в вертикальном расчленении биоценоза, ограниченная в пространстве (или во времени). Пространственно синузия может совпадать с горизонтом, пологом, слоем, ярусом биогеоценоза. Например, в сосновом лесу можно выделить синузию сосны, синузию брусники, синузию зеленых мхов и т. д. Парцелла - структурная часть в горизонтальном расчленении биоценоза, отличающаяся от других частей составом и свойствами компонентов. Парцеллу выделяют (ограничивают) по ведущему элементу растительности. Например, участки широколиственных деревьев в хвойном лесу. Самостоятельная работа № 4 Этапы эволюции биосферы Возникновение биосферы теснейшим образом связано с появлением жизни на Земле. Некоторые исследователи связывают появление жизни на нашей планете со временем ее остывания и оценивают возраст Земли по этому показателю в 24 млн. лет. Эти вычисления в 1861 г. сделал английский физик В. Томсон. Значительно позже, после открытия явления радиоактивности и разработки на ее основе метода измерения геологического времени, оказалось, что расчеты В. Томсона ошибочны. По современным представлениям, возраст Земли оценивается в 4,55 млрд лет, а сохранившиеся древнейшие участки земной коры — в 4 млрд. пет. Авторы большинства гипотез о происхождении жизни на Земле допускали, что наша планета в течение огромного промежутка времени была безжизненной и на ее поверхности происходила химическая эволюция, которая предшествовала биологической. На поверхности планеты, по их мнению, происходил медленный абиогенный синтез органических соединений, который в конечном счете привел к появлению примитивных форм жизни. Однако имеются и другие представления, согласно которым за такой период невозможно образование столь огромного количества видов живых организмов, то есть продолжительность существования жизни на Земле не укладывается в такие временные рамки. Анализ новых данных космохимии свидетельствует о раннем зарождении жизни в пределах Солнечной системы. Химическая эволюция вещества Земли и других планет, вероятно, предшествовала их образованию. Первичная атмосфера нашей планеты в основном была представлена углекислым газом. Но это химическое соединение не может самопроизвольно образовывать более сложные органические соединения. В настоящие время существует много подтверждений в пользу того, что зарождение жизни, возможно, произошло в космических условиях. Анализ вещественного состава метеоритов показывает, что содержащееся в них органическое вещество имеет достаточно сложный состав. Впервые органическое вещество в метеоритах обнаружил шведский химик И. Берцелиус в 1834 г. при анализе углистого хондритового метеорита Алаис. В органическом веществе метеоритов были обнаружены углеводороды, среди которых наиболее распространены соединения с 16 атомами углерода в молекуле, а также спирты, карбониловые соединения, аминокислоты и др. Следует отметить, что характерной особенностью органических соединений, обнаруженных в метеоритах, является отсутствие оптической плотности. Это свидетельствует об их происхождении за пределами Земли. Теоретические и экспериментальные данные, полученные в последнее время, позволяют сделать вывод: синтез относительно сложных органических соединений, предшествующих появлению живого вещества, — закономерный этап химической эволюции Солнечной системы. Эти органические вещества, образовавшиеся в космических условиях, вошли в состав многих тел, но на Земле реализовались возможности дальнейшей эволюции, что обеспечило возникновение саморегулирующихся высокомолекулярных систем — непосредственных предков первых живых организмов. Рассматриваются два варианта событий. Либо химическая эволюция, начавшаяся в космических условиях, продолжила свое развитие на Земле и в относительно короткое время привела к появлению первых живых организмов, либо образование молекул ДНК произошло в космических условиях, а реализация ее возможностей — в первых водоемах планеты, которые содержали некоторое количество органического вещества. Дальше в ходе геологической истории эволюция биосферы происходила по пути разрешения противоречия между безграничной способностью организмов к размножению и ограниченностью ресурсов, доступных в определенную геологическую эпоху. Данное противоречие разрешается путем овладения организмами новыми источниками вещества и энергии за счет приобретения ими новых качеств. В этом случае наследственная изменчивость является предпосылкой развития, а естественный отбор служит механизмом закрепления новых качеств. Переломным этапом в эволюции древней биосферы был переход от гетеротрофного режима питания к автотрофному, основанному на фотосинтезе. С появлением фотосинтезирующих организмов началось образование свободного кислорода, что со временем стало предпосылкой для создания в атмосфере озонового экрана. Это произошло около 4 млрд. лет тому назад. Увеличение содержания кислорода в атмосфере способствовало выходу органического мира на поверхность континентов. О времени появления живых организмов на суше точных палеонтологических данных нет. Переход от прокариотной флоры к эукариотной в древних морях происходил медленно, и обе группы длительное время существовали совместно. Соотношение между этими группами организмов постоянно изменялось и к настоящему времени оно сложилось в пользу эукариотов. Эволюция растений длительное время происходила в водной среде. Далее по мере накопления кислорода создались предпосылки для появления озонового экрана, который защищает все живое от ультрафиолетового излучения. Это в конечном счете создало условия для выхода растений из водной среды на континенты. Считается, что первыми растениями, появившимся на суше, были псилофиты — споровые низкорослые растения, напоминающие плауны. Потом псилофиты уступили место папоротникообразным растениям, которые в свою очередь сменились хвойными. Развитие растений, видимо, создало предпосылки для появления животных. Они так же, как и растения, произошли от одноклеточных организмов. В результате дифференциации функций отдельных клеток на определенном этапе эволюции образовались организмы, давшие затем начало многоклеточным. На основании палеонтологических данных в ходе эволюции органического мира выделяют ряд закономерностей:
- ускорение биологической эволюции в ходе геологического времени; - закономерность, впервые отмеченная В.О. Ковалевским в 1871 г.: «..каждая следующая большая эпоха Земли короче предыдущей, и в это короткое время успевало народиться и вымереть больше разнообразных форм, чем в предыдущую эпоху...» (цит. по Г.В. Войткевичу, В.А. Вронскому, 1996); - эволюция различных групп организмов протекала с разной скоростью; - существуют консервативные группы организмов, которые почти не изменились в ходе геологического времени (микроорганизмы, папоротниковые, плауны, голосеменные), однако они составляют небольшую часть из общего числа видов; - на фоне общей тенденции ускорения эволюции определенные эпохи отличались повышенным видообразованием. Вероятно, это было связано с радиоактивностью. С.Г. Неручев (1983) в геологической истории Земли выделил 30 эпох уранонакопления. Эти эпохи отличаются значительным усилением мутационных процессов, видообразования и сменой фауны и флоры; - среди животных в ходе геологического времени происходит направленное изменение нервной системы. На основании данных палеонтологии, геохимии и космохимии эволюцию биосферы Земли можно представить в виде трех последовательно сменяющихся этапов. Первый этап— восстановительный. Он начался в космических условиях и завершился появлением на Земле первой гетеротрофной биосферы. На этом этапе протекали каталитические и радиохимические реакции синтеза сложных органических соединений, отсутствовал свободный кислород, основным источником живых организмов была радиация. Этот период, вероятно, был коротким по времени. Второй этап — слабоокислительный — характеризовался появлением фотосинтеза. Он длился более 2 млрд. лет и закончился около 1,8 млрд. лет тому назад. Свободного кислорода образовывалось еще мало, и атмосфера состояла преимущественно из углекислого газа. Развитие организмов ограничивалось прокариотами. Третий этап — окислительный — связан с появлением фотоавтотрофной биосферы. Он начался с медленного роста содержания кислорода в атмосфере и завершился значительным ускорением эволюции организмов. Увеличение продукции кислорода привело к появлению растительного покрова и животных на континентах, что резко увеличило продукцию фотосинтеза. Под воздействием живого вещества сформировался современный химический состав атмосферы и растворенного вещества гидросферы. Биологическая эволюция, будучи необратимым процессом, предопределила необратимость эволюции биосферы в целом и создала предпосылки для ее перехода в качественно новое состояние — ноосферу, или сферу разума, когда все происходящие в биосфере изменения контролируются человеком.
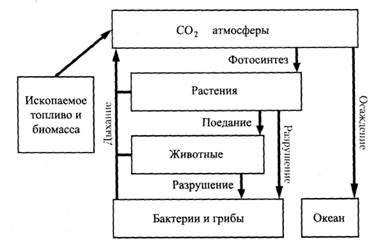
Самостоятельная работа № 5 Круговорот углерода Это один из самых важнейших биосферных круговоротов, поскольку углерод составляет основу органических веществ. В круговороте особенно велика роль диоксида углерода (рис.1). Запасы «живого» углерода в составе организмов суши и океана составляют, по разным данным, 550—750 Гт (1 Гт = 1 млрд. т), причем 99,5% его сосредоточено на суше, остальное - в океане. Кроме того, в океане содержится до 700 Гт углерода в составе растворенного органического вещества. Запасы неорганического углерода значительно больше. Над каждым квадратным метром суши и океана находится 1 кг углерода атмосферы и под каждым квадратным метром океана на глубине 4 км - 100 кг углерода в форме карбонатов и бикарбонатов. Еще больше запасов углерода в осадочных породах — в известняках содержатся карбонаты, в сланцах - керогены и т.д.
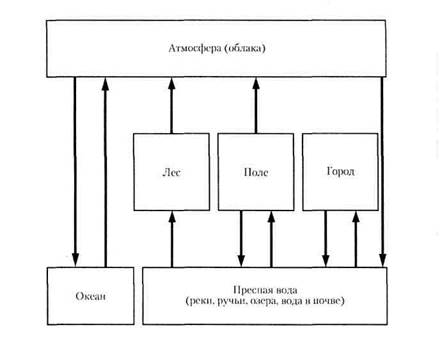
Рис.1. Круговорот углерода в биосфере Примерно 1/3 «живого» углерода (около 200 Гт) циркулирует, т. е. ежегодно усваивается организмами в процессе фото синтеза и возвращается обратно в атмосферу, причем вклад океана и суши в этот процесс примерно сходный. Несмотря на то, что биомасса океана много меньше биомассы суши, его биологическая продукция создается множеством поколений краткоживущих водорослей (соотношение биомассы и биологической продукции в океане примерно такое же, как в пресноводной экосистеме. До 50% (по некоторым данным — до 90%) углерода в форме диоксида возвращают в атмосферу микроорганизмы-редуценты почвы. В этот процесс равный вклад вносят бактерии и грибы. Возврат диоксида углерода при дыхании всех прочих организмов, таким образом, меньше, чем при деятельности редуцентов. Некоторые бактерии кроме диоксида углерода образуют метан. Выделение метана из почвы возрастает при переувлажнении, когда создаются анаэробные условия, благоприятные для деятельности метанобразующих бактерий. По этой причине резко увеличивается выделение метана лесной почвой, если древостой вырублен и вследствие уменьшения транспирации происходит ее заболачивание. Много метана выделяют рисовые поля и домашний скот. В настоящее время отмечается нарушение круговорота углерода в связи со сжиганием значительного количества ископаемых углеродистых энергоносителей, а также дегумификацией пахотных почв и осушением болот. В целом содержание диоксида углерода в атмосфере ежегодно увеличивается на 0,6%. Еще быстрее возрастает содержание метана - на 1-2%. Эти газы являются главными виновниками усиления парникового эффекта, который на 50% зависит от диоксида углерода и на 33% - от метана. Последствия усиления парникового эффекта для биосферы неясны, наиболее вероятный прогноз - потепление климата. Однако поскольку «машинами» климата являются морские течения, то вследствие их изменения при таянии ледников в ряде районов возможно существенное похолодание (в том числе в Европе в результате изменения течения Гольфстрим). Под влиянием изменения концентрации диоксида углерода значительно учащаются крупные стихийные бедствия (наводнения, засухи и т.д.) Приведенные данные характеризуют биогенный круговорот углерода. В круговороте участвуют и геохимические процессы, при которых происходит обмен атмосферного углерода и углерода, содержащегося в горных породах. Однако данных о скорости этих процессов нет. Полагают лишь, что их интенсивность менялась в истории планеты и парниковый эффект, который наблюдается сегодня, многократно проявлялся в прошлом при усилении геохимических процессов с выделением диоксида углерода и при ослаблении процессов, которые «оттягивали» его из атмосферы. Для того чтобы вернуть круговороту углерода равновесие, необходимо увеличить площадь лесов и сократить выброс газов при сжигании углеродистых энергоносителей. Круговорот воды Вода испаряется не только с поверхности водоемов и почв, но и живых организмов, ткани которых на 70 % состоят из воды (рис.2). Большое количество воды (около 1/3 всей воды осадков) испаряется растениями, особенно деревьями: на созидание 1 кг органического вещества в разных районах они расходуют от 200 до 700 л воды. Различные фракции воды гидросферы участвуют в круговороте по-разному и с разной скоростью. Так, полное обновление воды в составе ледников происходит за 8 тыс. лет, подземных вод - за 5 тыс. лет, океана - за 3 тыс. лет, почвы - за 1 год. Пары атмосферы и речные воды полностью обновляются за 10-12 суток. До развития цивилизации круговорот воды был равновесным, однако в последние десятилетия вмешательство человека нарушает этот цикл. В частности, уменьшается испарение воды лесами ввиду сокращения их площади и, напротив, увеличивается испарение с поверхности почвы при орошении сельскохозяйственных культур. Испарение воды с поверхности океана уменьшается вследствие появления на ее значительной части пленки нефти. Влияет на круговорот воды потепление климата, вызываемое парниковым эффектом. При усилении этих тенденций могут произойти существенные изменения круговорота, опасные для биосферы. Важную роль в годовом водном балансе биосферы играет океан. Испарение с его поверхности примерно в 2 раза больше, чем с поверхности суши.
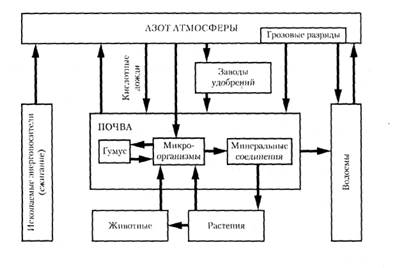
Рис.2 Круговорот воды в биосфере Круговорот азота Циркуляция азота в биосфере протекает по следующей схеме (рис.3): - перевод инертного азота атмосферы в доступные для растений формы (биологическая азотфиксация, образование аммиака при грозовых разрядах, производство азотных удобрений на заводах); - усвоение азота растениями; - переход части азота из растений в ткани животных; накопление азота в детрите; - разложение детрита микроорганизмами-редуцентами вплоть до восстановления молекулярного азота, который возвращается в атмосферу. В морских экосистемах азотфиксаторами являются цианобактерии, связывающие азот в аммиак, который усваивается фитопланктоном. В настоящее время вследствие уменьшения доли естественных экосистем биологическая азотфиксация стала меньше промышленной фиксации азота (соответственно 90-130 и 140 млн.т. в год), причем к 2020 г. ожидается увеличение промышленной азотфиксации на 60%. До половины азота, вносимого на поля, вымывается в грунтовые воды, озера, реки и вызывает эвтрофикацию водоемов.
Рис.3. Круговорот азота в биосфере Значительное количество азота в форме оксидов азота поступает в атмосферу, а затем в почву и водоемы в результате ее загрязнения промышленностью и транспортом (кислотные дожди). Этот азот был изъят из атмосферы экосистемами геологического прошлого и длительное время находился «на депоненте» в угле, газе, нефти, при сжигании которых он возвращается в круговорот. Например, в США азота с атмосферными осадками в год выпадает 20—50 кг/га, а в отдельных районах эмиссия достигает 115 кг/га. Экологически безопасной считается величина эмиссии азота 10—30 кг/га в год. При более высоких нагрузках происходят значительные изменения в экосистемах: почвы подкисляются, происходит выщелачивание питательных элементов в глубокие горизонты, возможно усыхание древостоев и массовое развитие заносных видов-нитрофилов. Кроме того, высокое содержание азота в растениях, выросших на загрязненных азотом почвах, повышает их поедаемость, что может привести к выпадению из растительных сообществ даже доминантных видов. Так, в некоторых пустошах Западной Европы после того, как в вереске повысилось содержание азота, массово размножился вересковый жук (его количество достигало 2000 экземпляров на 1 м2). Жук практически полностью выел этот кустарник из сообществ. Те же изменения в составе загрязняемых промышленным азотом сообществ отмечены и в Калифорнии. Однако не всегда кислотные дожди оказывают пагубное влияние на экосистемы. Экосистемы степной зоны, где почвы имеют слабощелочную реакцию, от выпадения кислотных дождей не только не страдают, но даже увеличивают свою продуктивность за счет дополнительного азота. Восстановление естественного круговорота азота возможно за счет уменьшения производства азотных удобрений, резкого сокращения промышленных выбросов оксидов азота в атмосферу и расширения площади посевов бобовых, которые симбиотически связаны с бактериями-азотфиксаторами. Круговорот кислорода Кислород атмосферы имеет биогенное происхождение, и его циркуляция в биосфере осуществляется путем пополнения запасов в атмосфере в результате фотосинтеза растений и поглощения при дыхании организмов и сжигании топлива в хозяйстве человека (рис.4). Кроме того, некоторое количество кислорода образуется в верхних слоях атмосферы при диссоциации воды и разрушении озона под действием ультрафиолетового излучения; часть кислорода расходуется на окислительные процессы в земной коре, при вулканических извержениях и др. Этот круговорот очень сложный, так как кислород вступает в разнообразные реакции и входит в состав очень большого числа органических и неорганических соединений, и замедленный. Для полного обновления всего кислорода атмосферы требуется около 2 тысяч лет (для сравнения: ежегодно обновляется около 1/3 диоксида углерода атмосферы). В настоящее время поддерживается равновесный круговорот кислорода, хотя в крупных густонаселенных городах с большим количеством транспорта и промышленных предприятий возникают локальные нарушения.
Рис.4. Круговорот кислорода в биосфере Однако отмечается ухудшение состояния озонового слоя и образование «озоновых дыр» (областей с пониженным содержанием озона) над полюсами Земли, что представляет экологическую опасность. Временные «дыры» возникают также над обширными районами вне полюсов (в том числе и над континентальными районами России). Причиной этих явлений является попадание в озоновый слой хлора и оксидов азота, которые образуются в почве из минеральных удобрений при их разрушении микроорганизмами, а также содержатся в выхлопных газах автомобилей. Эти вещества разрушают озон с более высокой скоростью, чем он может образовываться из кислорода под влиянием ультрафиолетовых лучей. Сохранение озонового слоя — одна из глобальных задач мирового сообщества. Для прекращения разрушения озонового слоя и его восстановления необходимо отказаться от использования хлорсодержащих веществ - хлорфторуглеродов (фреонов), используемых в аэрозольных упаковках и холодильных установках. Необходимо также уменьшить количество выхлопных газов двигателей внутреннего сгорания и дозы азотных минеральных удобрений в сельском хозяйстве. Отрицательно сказывается на здоровье человека и чрезмерно низкое содержание озона в атмосфере. Круговорот фосфора О круговороте фосфора за обозримое время можно говорить лишь условно. Будучи гораздо тяжелее углерода, кислорода и азота, фосфор почти не образует летучих соединений — он стекает с суши в океан, а возвращается в основном при подъеме суши в ходе геологических преобразований. По этой причине круговорот фосфора называют «открытым» (рис.5).
Рис.5 Круговорот фосфора в биосфере Фосфор содержится в горных породах, откуда выщелачивается в почву и усваивается растениями, а затем по пищевым цепям переходит к животным. После разложения мертвых тел растений и животных не весь фосфор вовлекается в круговорот, часть его вымывается из почвы в водоемы (реки, озера, моря). Там фосфор оседает на дно и почти не возвращается на сушу, лишь небольшое количество его возвращается с выловленной человеком рыбой или с экскрементами птиц, питающихся рыбой. Скопления экскрементов морских птиц служили в недалеком прошлом источником ценнейшего органического удобрения - гуано, но в настоящее время ресурсы гуано практически исчерпаны. Отток фосфора с суши в океан усиливается вследствие возрастания поверхностного стока воды при уничтожении лесов, распашке почв и внесении фосфорных удобрений. Поскольку запасы фосфора на суше ограничены, а его возврат из океана проблематичен (хотя в настоящее время активно исследуются возможности его добычи со дна океана), в будущем в земледелии возможен острый дефицит фосфора, что вызовет снижение урожаев (в первую очередь зерна). Поэтому необходима экономия ресурсов фосфора. Самостоятельная работа № 6 Экосистемы суши Размещение по земной поверхности основных наземных биомов определяют два абиотических фактора — температура и количество осадков. Климат в разных районах земного шара неодинаков. Годовая сумма осадков меняется от 0 до 2500 мм и более. При этом они выпадают равномерно в течение года или их основная доля приходится на определенный период — влажный сезон. Среднегодовая температура также варьирует от отрицательных величин до 38°С. Температуры могут быть практически постоянными в течение всего года (у экватора) или меняться по сезонам. Тундры (в северном полушарии к северу от тайги). Климат очень холодный с полярным днем и полярной ночью, среднегодовая температура ниже —5°С. За несколько недель короткого лета земля оттаивает не более чем на один метр в глубину. Осадков менее 200—300 мм в год. Растительность: отсутствуют деревья, господствуют медленно растущие лишайники, мхи, злаки и осоки, стелющиеся или карликовые кустарники (брусника, черника, карликовая береза). Животный мир. крупные травоядные копытные (северный олень, мускусный бык), мелкие роющие млекопитающие (лемминги), хищники, приобретающие зимой маскирующую белую окраску (песец, рысь, горностай, полярная сова). В тундре коротким летом гнездится большое число перелетных птиц, среди них особенно много водоплавающих, которые питаются имеющимися здесь в изобилии насекомыми и пресноводными беспозвоночными. Почвы тундровые — бедные с малой мощностью над слоем вечной мерзлоты. Бореальные хвойные леса (тайга) (северные районы Европы, Азии и Северной Америки). Климат: долгая и холодная зима, много осадков выпадает в виде снега. Растительность: господствуют вечнозеленые хвойные леса (ель, пихта, сибирская кедровая сосна, лиственница, сосна) с мощной лесной подстилкой. Животный мир: крупные травоядные копытные (лось, северный олень), мелкие растительноядные млекопитающие (заяц-беляк, белка, грызуны), волк, рысь, лисица, черный медведь, гризли, росомаха, норка и другие хищники, многочисленные кровососущие насекомые во время короткого лета. Множество болот и озер. Почвы подзолистые и дерново-п
|
||||
|
|
Последнее изменение этой страницы: 2017-02-07; просмотров: 632; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.154.251 (0.015 с.) |







 - необратимость эволюции. Это положение в 1893 г. сформулировал бельгийский палеонтолог Л. Далло: организм не может вернуться хотя бы частично к предшествующему состоянию, которое было в ряду его предков;
- необратимость эволюции. Это положение в 1893 г. сформулировал бельгийский палеонтолог Л. Далло: организм не может вернуться хотя бы частично к предшествующему состоянию, которое было в ряду его предков;