

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Суточные и сезонные аспекты экосистем
Сложные межвидовые взаимоотношения, определяющие функциональную целостность экосистем, отличаются относительной «свободой» структурных связей между отдельными компонентами. Виды в составе конкретных биоценозов могут викариировать, т. е. замещаться сходными по биологии другими видами. В зависимости от динамики численности и биологической активности отдельных популяций меняется направленность и интенсивность потоков веществ и энергии. Нестабильность абиотической составляющей экосистем определяет колебательный характер состава и функциональных связей в биоценозах. Иными словами, биоценозы как открытые системы, отличающиеся существенной ролью вероятностных процессов в их функционировании, непрерывно изменяются и развиваются под влиянием многих эндогенных и экзогенных факторов. Динамичность - одно из фундаментальных свойств экосистем, отражающее не только зависимость их от комплекса факторов, но и адаптивный ответ системы в целом на их воздействие.
Масштабы времени, в которых выражается динамика экосистем, различны: изменения в биоценозах могут иметь суточную или сезонную ритмику, длиться на протяжении ряда лет или же охватывать целые геологические эпохи, отражая развитие биосферы в целом.
Изменения облика биоценозов, связанные с динамикой активности, сезонными ритмами в жизни растений и животных, сменами видового состава на протяжении годового цикла, отражают комплекс приспособлений экосистемы в целом к суточной и сезонной динамике условий существования. Характерной чертой такого типа динамики является сохранение принципиальных свойств данной экосистемы, поддержание ее целостности и функциональной устойчивости. Даже сезонные изменения видового состава входят в общую характеристику каждой данной экосистемы, поскольку закономерно повторяются год за годом.
Суточные аспекты биоценозов. При закономерных ритмичных изменениях в масштабах суток не происходит принципиальных перестроек видового состава и основных форм взаимоотношении в биоценотической системе. Подчеркивая это, обычно говорят не о суточной динамике, а о суточных аспектах биоценоза. Смена аспектов в этом масштабе времени определяется характером активности тех видов, которые отличаются отчетливой суточной ритмикой жизнедеятельности. Так, в лесах умеренной зоны в дневном аспекте биоценоза господствуют насекомые, птицы и некоторые другие животные, отличающиеся дневной активностью: среди цветковых растений в период цветения красочный дневной аспект определяется тем, что большинство видов раскрывает цветки днем. В ночное время на первое место выходит активность ночных видов животных (ночные бабочки, многие млекопитающие, из птиц - козодои, совы и некоторые другие), а также растения, опыляемые ночными животными.
Среди рыб также имеются формы с дневной или ночной активностью. Известны суточные вертикальные миграции планктона и сопутствующие им перемещения ряда планктоноядных животных. Число примеров можно было бы увеличить.
В некоторых случаях суточные аспекты биоценозов определяются экзогенно. Так, в пустынях континентального климата днем - в самое жаркое время суток - наблюдается резкий спад активности большинства видов -даже тех из них, которые в принципе отличаются дневной активностью. Более того, в летний период, когда суточные изменения температуры наиболее экстремальны, ряд дневных видов меняет характер активности на сумеречную, а подчас и на ночную (некоторые насекомые, змеи и др.).
Суточные аспекты биоценозов отражают их «нишевую структуру». Разделение периодов активности во времени снижает уровень прямой конкуренции (интерференции) и таким образом открывает возможность сосуществования видов со сходными биологическими требованиями. Эксплуатационная конкуренция при этом в принципе сохраняется, но отличия во времени активности с неизбежностью влекут за собой некоторый «сдвиг» в пищевых спектрах, а стало быть, ослабление конкурентных связей и в этом аспекте. В целом расхождение в суточной активности приводит к усложнению биоценоза, повышению биологическою разнообразия и более полному использованию ресурсов среды.
Сезонные аспекты биоценозов. Сезонные изменения затрагивают более фундаментальные характеристики экосистем. В первую очередь это касается видового состава биоценозов. В неблагоприятные сезоны года ряд видов мигрируют в районы с лучшими условиями существования. Такое явление хорошо известно для перелетных птиц. ряда видов копытных млекопитающих и др.. Оседлые виды составляют основное ядро биоценоза, тогда как сезонные определяют его облик, а также характер биоценотических связей в отдельные периоды
Некоторые виды отличаются «пульсирующим» характером сезонной активности. Они переносят неблагоприятные периоды в состоянии заторможенной жизнедеятельности (диапауза насекомых, оцепенение пойкилотермных и спячка гомойотермных животных, инцистирование простейших и т. п.).
Растительные сообщества также меняются по сезонам как структурно (листопад, выпадение однолетников, засыхание травянистой растительности), так и функционально (изменение интенсивности фотосинтеза, накопления биомассы и пр.).
Во всех случаях уменьшение числа активных видов влечет за собой снижение общего уровня биогенного круговорота веществ. Таким образом, такой «формальный» параметр, как число видов, оказывается важным фактором регуляции биосферных процессов.
Сезонные аспекты биоценозов лучше всего выражены в ландшафтно-климатических зонах, отличающихся резкими изменениями физических параметров среды летом и зимой. В частности, они хорошо выражены в тундрах, для которых в летнее время характерен большой набор видов птиц, насекомых и других животных; на зиму большая часть из них мигрирует на юг (большинство птиц, северные олени), а другие впадают в оцепенение и выключаются из активной жизни (насекомые, другие беспозвоночные). Длинная полярная ночь, закрывающая возможность фотосинтеза, еще более усугубляет функциональную значимость сезонной динамики тундровых экосистем.
В тропиках сезонность функционирования биоценозов выражена не столь ритмично, но все же имеет место. Наиболее распространенная ее форма - смена засушливых и влажных периодов - явление не столь ритмичное, как смена летних и зимних аспектов, но биологически не менее значимое.
Сезонные биологические процессы в водной среде связаны с гидрологическими сезонами. Они не совпадают в различных зонах и характеризуются закономерными колебаниями видового состава водных организмов, их биомассы и биологической активности. Так, морские планктонные сообщества демонстрируют отчетливую смену фаз развития во времени. «Биологическая весна» характеризуется массовым развитием фитопланктона («период цветения моря»), тогда как большинство видов зоопланктона на этом фоне начинает размножение. «Биологическое лето», напротив, отличается массовым развитием зоопланктона и уменьшением биомассы планктонных водорослей. «Биологическая осень» в арктических морях знаменуется общим уменьшением планктона, а в морях умеренной зоны в это время наблюдается «осеннее цветение» - вторая вспышка численности планктонных организмов. Зима - время минимума биомассы планктона, причем многие виды проводят этот сезон в покоящихся стадиях. Длительность отдельных сезонов широко колеблется, в тропиках вегетационный сезон длится практически круглый год, а и арктических морях - обычно не более 2-3 месяцев.
Суточные и сезонные аспекты биоценозов хотя и могут быть связаны с изменениями числа видов, но не затрагивают принципиальных параметров данного типа биоценоза. В этих случаях можно говорить о количественных изменениях, не меняющих качественных характеристик системы. В ряде случаев динамика биоценозом может иметь принципиально иной характер, когда под влиянием комплекса факторов меняются фундаментальные свойства экосистемы, и динамика их выражена серией сменяющих друг друга сообществ.
ЭКОЛОГИЧЕСКИЕ СУКЦЕССИИ
Развитие биоценозов, при котором имеет место замещение во времени одного сообщества другим, называют экологической сукцессией. В большинстве случаев процессы сукцессии занимают временные промежутки, измеряемые годами и десятилетиями, хотя и отдельных случаях смены сообществ следуют с большей скоростью (например, во временных водоемах). Наряду с этим известны вековые изменения экосистем, отражающие общие пути эволюции биосферы.
Общие понятия о сукцессиях. Разработка проблемы сукцессий началась в ботанике, и по сей день основные положения этой концепции базируются на изучении фитоценозов. Это определяется не только историческими причинами, но и тем, что смены сообществ базируются на функциях автотрофов. Гетеротрофная составляющая биоценозов формируется на базе фитоценоза и лишь вторично начинает влиять на его состав и свойства.
Впервые динамику сообществ описал, по-видимому, Варминг. Но наиболее существенный вклад в разработку концепции сукцессий сделали американские ботаники Коулес и особенно Клементс. Основная позиция Ф. Клементса заключается в том, что изменения во времени - естественное свойство экологических сообществ. Первопричиной смены фитоценозов он считал изменение отдельных климатических факторов или их комплекса, а реакция экосистем в виде смены последовательного ряда сообществ представляет адаптивный ответ на экосистемном уровне. Сукцессия, по Ф. Клементсу, завершается формированием сообщества, наиболее адаптированного по отношению к комплексу климатических условий. Такое сообщество он называй «климакс-формация», или просто климакс.
Таким образом, концепция климакса подразумевает, что в пролетах региона, характеризующегося более или менее однородным климатом фитоценозы, завершившие сукцессионный процесс, образуют климаксовое сообщество независимо от того, с какого типа начиналась сукцессия. Смены растительности, начинающиеся от разных сообществ и заканчивающиеся климаксом, называют сукцессионными сериями. Сукцессионные серии в зависимости от условия влажности делят на гидросерии (исходными являются сообщества влажных местообитаний) и ксеросерии (начинаются от сухих сообществ). Процесс сукцессии меняет их на ассоциации промежуточных по влажности местообитаний (мезосерии), которые существуют в динамическом равновесии с региональным климатом. Ф. Клементс полагал, что все серии сукцессионных смен сообществ могут быть только прогрессивными.
Современная концепция экологических сукцессий расходится с представлениями Ф. Клементса лишь по некоторым относительно второстепенным пунктам. Наиболее принципиальный из них заключается в том, что климакс как завершающая формация есть тоже лишь временное состояние; в процессе вековых изменений климата и других свойств среды имеют место «крупномасштабные» изменения экосистем. Кроме того, отрицается жесткая трактовка гипотезы «моноклимакса». По Ф. Клементсу, в одной биоклиматической зоне возможен лишь один вариант климаксового сообщества, современные исследования показали, что многообразие влияний на ход сукцессии приводит к тому, что устойчивым (равновесным) в данных географических условиях могут быть несколько типов биоценозов (гипотеза «поликлимакса») или даже мозаика таких сообществ. Наконец, признано, что в определенных условиях сукцессия может быть регрессивной, направленной на обеднение и упрощение сообществ. Особенно часто сукцессии такого рода (дигрессии) возникают в результате антропогенных воздействий на биоценоз, нарушающих оптимальность условий существования тех или иных видов.
Не выдержало испытания временем и утверждение Ф. Клементса об исключительном значении климата как движущей силы сукцессий. Смены сообществ могут происходить и под влиянием других факторов - таких, как изменения рельефа, почвы, гидрологического режима и т. и. Важнейшее значение в современной экологии придается биоценотическим факторам сукцессии: виды растений (а также и животных), участвующие в сукцессионных сообществах, изменяют условия обитания для других видов, таким образом «подготавливая почву» для последующего этапа сукцессии.
В соответствии с этим в современной экологии различают сукцессии экзоэкогенетические (или аллогенные) и эндоэкогенетические е (автогенные). В первом случае речь идет о сукцессионных сменах, вызванных внешними, абиотическими причинами. Примеры экзоэкогенетических сукцессий можно найти в различных воздействиях на биоценозы со стороны человека; мелиоративное осушение болот, загрязнение водоемов, неумеренный выпас скота и т. и. Эндоэкогенетические сукцессии вызываются в первую очередь изменением структуры и системы связей в существующих сообществах. Впрочем, эти две категории сукцессий взаимосвязаны и могут переходить одна в другую; об этом создатель теории биогеоценологии акад. В.Н. Сукачев писал еще в первой половине текущего столетия.
Начальный этап сукцессий В.Н. Сукачев называл этапом сингенеза Он определил это понятие как «процесс первоначального формирования растительного покрова, связанный с вселением (миграцией) растений на данную территорию, их отбором в процессе приспособления к ее условиям (эцезисом), затем и конкуренцией между ними из-за средств жизни». Сингенетическая сукцессия, направляемая исходно исключительно абиотическими свойствами местности, приводит к формированию фитоценоза, после чего сукцессия идет по принципу эндоэкогенетической, приводя к устойчивому сообществу, соответствующему понятию климакса.
В наиболее общем виде сукцессии, по Ф. Клементсу, проходят через фазы обнажения (появление незаселенного пространства), миграции (заселение его первыми, пионерными формами жизни), эцезиса (колонизация и приспособление к конкретным условиям среды), соревнования (конкуренция с вытеснением ряда первичных вселенцев), реакции (обратное воздействие сообщества на биотоп и условия существования) и, наконец, стабилизации (формирование климаксового биоценоза). Таким образом, сукцессионные серии начинаются, как правило, с экзоэкогенеза, переходящего в эндоэкогенез по мере формирования внутренних связей в возникающем фитоценозе.
Важным механизмом стабилизации В.Н. Сукачев считал конкурентные отношения, в процессе которых достигается равновесное состояние, характеризующее завершающее сообщество. Более подробная классификация механизмов сукцессии предусматривает три категории отношений между организмами в сукцессионных сериях. Модель облегчения или стимуляции соответствует фазе эндоэкогенетической сукцессии и заключается в том, что ранние поселенцы своей деятельностью изменяют среду, делая ее доступной для следующей волны колонистов. Модель толерантности проявляется в виде конкурентных отношений, в результате которых происходит отбор более толерантных и конкурентоспособных видов. Смена видов основывается на их различии в стратегии потребления ресурсов; виды более поздних стадий более устойчивы. По модели ингибирования все виды сообщества способны одновременно колонизировать открывшееся местообитание, устойчивы к вторжению конкурентов, но более поздние вселенцы способны закрепиться и увеличить численность только после выпадения кого-либо из предшественников. Так, в тропических дождевых лесах ползучий кустарнике Lantana занимает большие площади, практически исключая поселение на них деревьев. В прериях сукцессионная стадия однолетников длится до 15 лет, прежде чем они будут замещены многолетниками. По этой модели изменение среды предшественниками делает ее менее пригодной для последующих вселенцев.
Виды растений, входящие в состав первичных и последующих сообществ сукцессионной серии, могут отличаться по эколого-физиологическим свойствам. Показано, в частности, что раннесукцессионные виды травянистых и древесных растений отличаются большей скоростью фотосинтеза. Так, незимующие однолетники потребляют 38-18 мл СО2 /дм2 ч, озимые однолетники — 22-20, раннесукцессионные деревья — 26-11, позднесукцессионные деревья — 18-6 мл СО2 /(дм2 ч).
При этом сукцессия может проходить как на базе К-стратегии (отбор на признаки, способствующие закреплению в сукцессионном ряду), так и по линии r-стратегии (эффективные механизмы избегания сукцессионного пресса, поиски и захват новых мест, где сукцессия только началась).
Первичные и вторичные сукцессии. По общему характеру сукцессии подразделяются на первичные и вторичные. Первичные сукцессии начинаются на cyбстрате, не измененном (или почти не измененном) деятельностью живых организмов. Так, через серию промежуточных сообществ формируются устойчивые биоценозы на скалах, песках, обрывах и т. п. Такие сукцессии относятся к категории ксеросерий и называются ксерическими сукцессиями. Одна из основных функций сукцессий такого рода - создание (или изменение) почвы первичными колонистами.
Сукцессии, формирующие экосистемы на местах, исходно не заселенных называют экогенетическими. Формирование скальных биоценозов - классический пример таких сукцессий. Процесс начинается с поселения на скалах накипных лишайников. Уже на этой стадии формируется комплекс видов микроскопических водорослей, простейших, нематод, некоторых насекомых и клещей, который способствует созданию первичной почвы. Позднее здесь поселяются другие формы лишайников, специализированные виды мхов; еще позднее - на базе возникшей почвы - вселяются сосудистые растения. Параллельно идет обогащение животного населения.
Сходным образом идет смена сообществ на ледниковых отложениях в виде очень тонкой, бедной биогенами почвы. Наблюдения на Аляске показали, что формирование фитоценоза начинается с мхов и осок; вслед за ними в сообщество включаются стелющиеся, а затем и кустарниковые формы ив. Позднее (примерно через 20-25 лет) возникают ольшатники; вслед за ними появляется ель, которая и составляет основу завершающего сообщества в виде смешанного леса, формирующегося примерно через 100 лет после начала сукцессии.
Гидрические сукцессии начинаются и открытых водах мелких отер, верховых болот, маршей. Постепенное зарастание водными растениями, идущее по краям открытого водоема, ведет к накоплению на дно детрита, образованию накоплений торфа и в конце концов к обмелению водоема. В других местах побережья развивается сплавина из сфагновых мхов и болотных сосудистых растений. Накопление растительной массы способствует образованию почвы. Обмеление вкупе с увеличением толщины сплавины приводит к превращению водоема в болото. Позднее здесь селятся кустарники и древесные породы, идет процесс усыхания болота и развивается лесная растительность. Изменение растительной части сообщества сопровождается сдвигами н фауне: обитатели водоема постепенно замещаются околоводными, а позднее - болотными и лесными видами.
В водной среде сукцессия такого рода прослежена экспериментально. На поверхности скалы в зоне прилива была очищена от макроорганизмов площадка среди мидиевого биоценоза. Первыми ее заселили водоросли. Вслед за ними вселились питающиеся водорослями животные, в том числе моллюск Aстаеа. В период размножения на площадку вселились мидии Mytilus и усоногие раки Mitella и Balanus. Эти сидячие формы постепенно расширили занятую площадь, делая местообитание непригодным для Астаеа эти моллюски переместились в более высокую зону, где мидии и усоногие существовать не могут. Поскольку в этом случае сформировался не новый биоценоз, а по существу восстановилось искусственно нарушенное сообщество, в данном случае правильнее говорить не об экогенетической, а о демутационной смене. Направленность такого рода более типична для вторичных сукцессий.
Вторичные сукцессии развиваются на субстрате, первоначально измененном деятельностью комплекса живых организмов. Такие сукцессии чаще всего имеют восстановительный (демутационный) характер.
Примером вторичных сукцессий демутационного типа может служить восстановление климаксового лесного биоценоза после пожаров (а в наше время и вырубок). В таежной зоне Евразии появление открытого пространства на месте еловых лесов в результате пожара или сплошной рубки коренным образом меняет режим освещения, температуры, влажности и других факторов. Изменения эти неблагоприятны для комплекса таежных видов растений и животных, их развитие угнетается. Зато на освещенных, относительно сухих и хорошо прогреваемых местах формируется временное одноярусное сообщество из светолюбивых трав. Первыми здесь развиваются виды с легко распространяемыми семенами: вейник. Иван-чай и др. - это стадия открытой вырубки. Позднее начинается лесовозобновление, прорастают светолюбивые лиственные породы (осина, береза, ивы и др.); поселяются кустарники. Одновременно формируется связанный с лугово-кустарниковой растительностью комплекс животного населения: многочисленные и разнообразные насекомые и другие беспозвоночные, грызуны, исходно-связанные с лугами и лесными полянами, развитие ягодных кустарников привлекает большое число видов птиц, наличие хорошо прогреваемых мест - рептилий и т. д. Формируется богатое и разнообразное сообщество зарастающей вырубки.
Эта стадия занимает в среднем 2 - 3 года, после чего начинается интенсивное развитие светолюбивых мелколиственных древесных пород (осины, березы). Постепенно подросшие деревья вытесняют кустарники и наиболее светолюбивые виды трав; кустарниково-луговое сообщество сменяется лиственным жердняком - молодым лесом с несомкнутыми кронами. Это влечет за собой и изменение животного населения на стадии жердняков оно обедняется за счет эмиграции милов, ранее связанных с кустарниками и богатым разнотравьем. После смыкания Крон и перехода сообщества от стадии жердняка в фазу лиственною леса (через 10-15 лет от начала сукцессии) биоценоз вновь несколько усложняется за счет появления более или менее полного комплекса растительности и животного населения, характеризующего спелые лиственные леса. Под древесным пологом в условиях затенения и повышенной влажности начинается интенсивное прорастание семян ели. Постепенно хвойный молодняк окончательно заглушает луговую травянистую растительность; ее сменяют мхи и лесное разнотравье. Улучшение условий для роста ели ингибирует восстановление мелколиственных лесных пород. Старые лиственные деревья затеняются выходящими в первый ярус елями; смыкание еловых крон еще больше угнетает березы и осины, которые выступают уже в качестве второстепенных членов древостоя. В конце концов лиственные деревья выпадают, и восстанавливается исходный тал лесного сообщества с господством ели. Соответственно изменяется и фаунистический комплекс, прошедший в процессе сукцессии фазы сорно-луговой растительности, кустарников, лиственного жердняка и спелого леса. Весь процесс от вырубки (пожара) до формирования устойчивого таежного биоценоза занимает в среднем 90-150 лет.
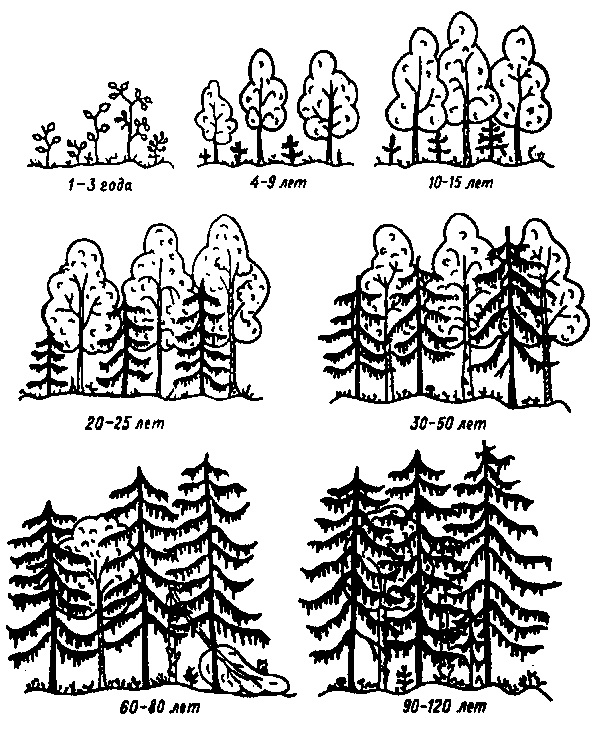
Рисунок 1. Схема возобновления елового леса после вырубки
К вторичным сукцессиям относятся и такие, при которых исходной силой, вызывающей смену сообществ, оказываются нарушения стабильных взаимодействий в биоценозе. Таковы, в частности, зоогенные сукцессии, выражающиеся в изменении фитоценоза при чрезмерном выпасе скота. Процесс перестройки и ксерофизации фитоценоза в результате перевыпаса сопровождается расселением и ростом численности сухолюбивых видов животных — насекомых, птиц, млекопитающих (суслики, тушканчики, степные виды полевок и др.). На пастбищах увеличивается число копрофагов, специфических паразитов домашнего скота, а также концентрируются крупные хищники и падальщики.
|




