Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Ядерно - цитоплазматическое взаимодействиеСодержание книги
Поиск на нашем сайте Процессы взаимодействия ядра и цитоплазмы обеспечивают морфофункциональное единство клетки. Из цитоплазмы в ядро поступают вещества (обычно белковой природы), которые регулируют активность генов. В ядре также находятся нуклеотиды, АТФ; ферменты, необходимые для репликации ДНК, синтеза РНК, а также белки, входящие в состав хроматина, ядрышек и других структур ядра. Из ядра в цитоплазму выходят продукты генной активности: и-РНК, т-РНК, субъединицы рыбосом (р-РНК в комплексе с белками), которые обеспечивают синтез белков в цитоплазме – структурных белков, белков-ферментов. Таким образом, ядро управляет всеми белковыми синтезами и через них функциональными и морфологическими процессами в клетке, а цитоплазма − регулирует (по принципу обратной связи) активность генетического аппарата ядра и снабжает его материально и энергией. Выделяют два состояния ядра: интерфазное и делящееся. В интерфазном ядре происходит реализация генетической информации, а в период деления ядра – ее передача. Структура интерфазного ядра В интерфазном ядре различают: ядерную оболоночку, кариоплазму (ядерный матрикс, нуклеоплазма), хроматин и ядрышко (одно или несколько) (рис. 6.1.).
Рис.6.1. Схема строения клеточного ядра Ядерная оболочка образована двумя мембранами (наружной и внутренней), между которыми находится перинуклеарное пространство шириной от 20 до 60 нм. Наружная мембрана в некоторых местах переходит в каналы эндоплазматической сети, на ней располагаются рибосомы. Изнутри ядерная оболочка подстелается белковой сеткой – ядерной ламиной. До ламины теломерными участками присоединяются хромосомы (нити хроматина). Ядерная оболочка пронизана множеством пор, диаметром около 90 нм, через которые происходит обмен веществ между ядром и цитоплазмой. Основная функция ядерной оболочки: регуляция потоков веществ между ядром и цитоплазмой. Через поры из ядра выходят разные виды РНК, субъединицы рибосом, а внутрь ядра поступают необходимые белки, нуклеотиды, ионы, вода. Кариоплазма (ядерный матрикс) – внутреннее содержимое ядра, в котором располагаются хроматин и одно или несколько ядрышек. В состав кариоплазмы входит вода, минеральные соли, белки-ферменты, нуклеотиды, РНК, ионы. В нуклеоплазме имеются белковые фибриллы толщиной 2-3 нм, выполняющие роль внутреннего скелета ядра и соединяющие ядрышка, нити хроматина, ядерные поры. Кариоплазма осуществляет взаимосвязь всех структур ядра и обменные процессы с цитоплазмой клетки. Генетический материал в интерфазном ядре находится в виде хроматина. Хроматин – комплекс ядерной ДНК с белками (дезоксирибонуклеопротеин – ДНП). В интерфазном ядре хромосомы деконденсированы (деспирализованы) и не выделяются по отдельности, а воспринимаются все вмести в виде глыбок, гранул, нитчатых структур, которые получили название хроматина. Существование хромосом в неделящихся клетках в форме хроматина обеспечивает возможность удвоения ДНК и реализации заключенной в ней генетической информации. Различные участки молекул ДНК в составе хроматина обладают разной степенью спирализации, а поэтому различаются степенью окраски и характером генной активности. После фиксации и специфической окраски в кариоплазме становятся видимыми два типа хроматина: хорошо окрашенный гетерохроматин и светлый эухроматин. Гетерохроматин – это сильно конденсированные (уплотненные), а поэтому функционально неактивные участки хромосом. До 90% хроматина находится именно в такой форме. На электронно-микроскопических фотографиях гетерохроматин выглядит как сильно окрашенные темные участки ядра, расположеные в основном вблизи внутренней ядерной мембраны и вокруг ядрышка. Эухроматин – слабо конденсированные (деспирализованные) функционально активные участки хромосом, расположеные между глыбами гетерохроматина. Под электронным микроскоп он выявляется в виде светлых участков ядра. На эухроматиновых участках считывается информация и образуется РНК. В клетках с интенсивным синтезом белка эухроматина больше. Считается, что гетерохроматин неактивен в связи с высокой конденсацией, а эухроматин – активен. С другой стороны, только незначительная часть генов эухроматина активна, т.е. нахождение участков хромосом в состоянии эухроматина является недостаточным условием для экспрессии генов. Различают конститутивный (структурный) и факультативный гетерохроматин. Конститутивный – гетерохроматин содержится в обеих гомологичных хромосомах и локализуется в околоцентромерных и теломерных участках хромосом, а также определенных, постоянно конденсованных, участках некоторых хромосом. Полагают, что его роль заключается в прикреплении хроматина к ядерной оболочке, взаимном узнавании гомологичных хромосом в мейозе, разделении соседних структурных генов, участии в процессах регуляции их активности. Факультативный гетерохроматинприсутствует только в одной из гомологичных хромосом. Примером хроматина такого типа может быть половой хроматин, который в норме выявляется в клетках гомогаметного пола. Половой хроматин – сильно спирализованная генетически инактивированная Х-хромосома, которая при специфическом окрашивании выявляется в интерфазных ядрах в виде темного тельца (тельца Барра). Количество телец полового хроматина в ядре на единицу меньше чем число Х-хромосом, т.е. равно nx-1(рис 6.2.).
Рис. 6.2. Половой хроматин (тельца Барра): а -при кариотипе ХХ, ХХУ; б -при кариотипе ХУ, ХО; в -при кариотипе ХХХ В сегментоядерных лейкоцитах Х-половой хроматин имеет вид специфической структуры, которая получила название «барабанные палочки» (рис. 6.3.)
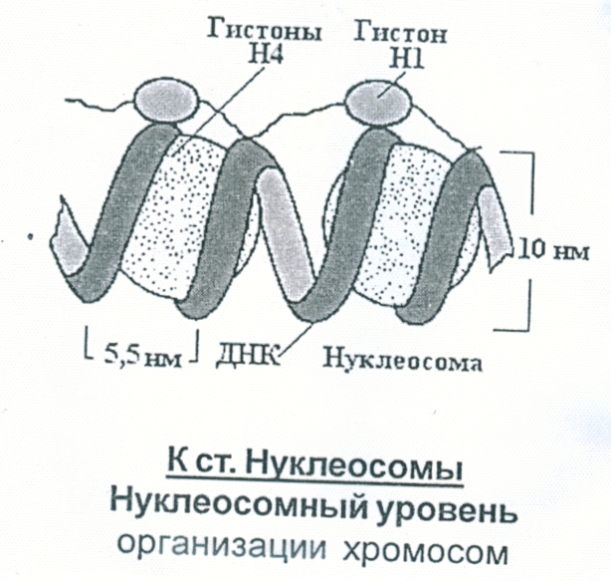
Рис.6.3. Барабанные палочки в сегментоядерных лейкоцитах при кариотипе ХХ Определение Х-полового хроматина используют для выявления нарушений кариотипа по числу Х-хромосом. Химическую основу хроматина составляет двуспиральная ДНК (около 40%) и беки-гистоны (40%). Известно 5 типов гистонов, связанных с ДНК: Н1, Н2А, Н2В, Н3 и Н4. Благодаря основным радикалам гистоны взаимодействуют с ДНК, а благодаря гидрофобным радикалам они взаимодействуют между собой. Эти взаимодействия приводят к образованию нуклеосом. Нуклеосома – нуклеопротеиновая структура, состоящая из гистонового октамера (глобула), обвитого участком ДНК длиной 140-160 пар нуклеотидов (рис. 6.4.).
Рис. 6.4. Нуклеосомы
Гистоновый октамер состоит из 8 молекул (по 2 молекулы гистонов Н2А, Н2В, Н3, Н4). Соседние нуклеосомы соединяются друг с другом короткими отрезками ДНК длиной 60 нуклеотидных пар. Молекула ДНК принимает участие в образовании большого количества нуклеосом. В результате образуется хроматиновое волокно в виде «бусинок на нитке», при этом происходит укорочение ДНК примерно в 7 раз (рис 6.5.).
Двойная спираль ДНК –
Нуклеосомный уровень –
Уровень фибрилл –
Уровень петельных доменов –
Хромонемный уровень –
Метафазная хромосома – Рис. 6.5. Уровни организации хроматина эукариот.
Нуклеосомный уровень организации хроматина свойствен как для эухроматина, так и для гетерохроматина. Но в тех локусах хроматина, на которых в данное время функционируют ферментные комплексы (репликации, транскрипции, репарации) ДНК, освобождается от взаимодействия с гистонами. В этих случаях нуклеосомная организация временно исчезает с тем, чтобы в последующим снова восстановится. В отличие от этого, в гетерохроматине до нуклеосомного уровня добавляются последующие уровни компактизации хроматина. Второй уровень – образование хроматиновых фибрил (глобул) за счет сближения нуклеосом, что обеспечивается гистоном Н1. Третий уровень – образование петельных доменов (хромомер) за счет укладки хроматиновых фибрилл в петли. Четвертый уровень – хромонемный. Образуется за счет сближения в линейном порядке хромомерных петель с образованием хромонемной нити. Таким образом, хромосомы образуются из хроматина интерфазного ядра в процессе спирализации и конденсации дезоксирибонуклеопротеинов. Хромосомы Хромосомы (греч. хрома – краска, сома - тело) были так названы в связи со способностью их к интенсивному окрашиванию. Являются еденицами организации генетического материала и обеспечивают его точное распределение при делении клетки. Хромосомы лучше всего различимы (и изучаются) на стадии метафазы. На стадии метафазы митоза хромосома состоит из двух нуклеопротеидных копий - хроматид. Хроматиды соединяются в области неспирализованой первичной перетяжки – центромеры. В центральной части центромеры расположен кинетохор, к которому присоеденяются нити веретена деления во время митоза. Центромера делит хромосому на два плеча. Плечи имеют свои обозначения: короткое – p, и длинное – q. В зависимости от расположения центромеры различают следующие типы хромосом: · метацентрические (равноплечие), в которых центромера расположена по средине (p=q); · субметацентрические (неравноплечие), когда центромера смещена к одному концу хромосомы (q>p); · акроцентрические (одноплечие или палочковидные), когда центромера расположена на краю хромосомы и одно плечо очень короткое (одноплечие - q). Некоторые хромосомы имеют вторичные перетяжки, отделяющие спутники или сателлиты (рис.6.5.). Они характерны, например, для хромосом 13 – 15, 21 и 22 человека. В области вторичной перетяжки ряда хромосом имеется ядрышковый организатор – область формирования ядрышка.
А.
Б. Рис. 6.5. Схема строения метафазной хромосомы (А), типы хромосом (Б). Концевые участки хромосом получили название теломеры. Они препятствуют соеденинею хромосом между собой и сохраняют хромосому как дискретную индивидуальную единицу. Правила хромосом: 1. Правило постоянства числа хромосом – у всех организмов одного вида соматические клетки имеют постоянное число хромосом. 2. Правило парности хромосом – в соматических клетках с диплоидным набором хромосом имеються пары одинаковых по структуре, форме и генному составу хромосом – так называемые гомологичные хромосомы (одна – материнская, вторая – отцовская). 3. Правило индивидуальности хромосом – каждая пара хромосом имеет свои особенности, что проявляется в размерах, форме, положении центромеры, чередовании светлых и темных полос при диференциальной окраске. 4. Правило непрырывности хромосом. В основе его лежит удвоение ДНК и формирование новых сестринских хроматид перед каждым делением клетки. Набор хромосом соматических клеток конкретного вида живых организмов, называют кариотипом. Количество хромосом в кариотипе не зависит от уровня организации живых организмов. Некоторые протисты имеют их более тысячи, у шимпанзе – 48, у собаки - 78, у коровы - 60, у дрозофилы – 8, у рака отшельника - 254 и т.д. У человека кариотип представлен 46 хромосомами (23 пары); 44 (22 пары) – аутосомы и две половые хромосомы: ХХ у женщины и ХУ у мужчины. Исследуется кариотип обачно на стадии метафазы, когда каждая хромосома состоит из двух хроматид и максимально спирализована. Для изготовления препаратов хромосом человека обычно используют клетки лейкоцитов переферической крови. Сначала до культуры клеток добавляют колхицын, который останавливают деление клеток на стадии метафазы. Далее прибавляют гипотонический раствор 0,56% NaCl который отделяет хромосомы одну от другой, после чего их фиксируют и окрашивают. После такой обработки каждая хромосома имеет четкие очертания и видима в световом микроскопе (рис. 6.6)
Рис. 6.6. Кариотип и идиограмма хромосом человека
Для того что бы провести анализ хромосом, образующих каріотип, их располагают в виде идиограммы. Идиограмма – это систематизированный кариотип, в котором хромосомы располагаются попарно в порядке уменьшения их величины. Исключение делается для половых хромосом, которые располагаются последними. Точно расположить хромосомы по величине сложно, так как некоторые пары хромосом весьма незначительно отличаются по размерам. В связи с этим в 1960 г была предложена Денверская классификация хромосом. По этой классификации, кроме размеров хромосом, учитывают их форму, положение центромеры, наличие вторичных перетяжек и спутников. 23 пары хромосом разбиты на 7 групп от А до G. Важной характеристикой хромосом является центромерный индекс (ЦИ), который отражает отношение длины короткого плеча к длине всей хромосомы (в %).
|
||
|
|
Последнее изменение этой страницы: 2016-12-27; просмотров: 2605; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.96 (0.008 с.) |