Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Пути передачи сигнала клеткеСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Сигнальная молекула
Клетка-продуцент сигнальной молекулы
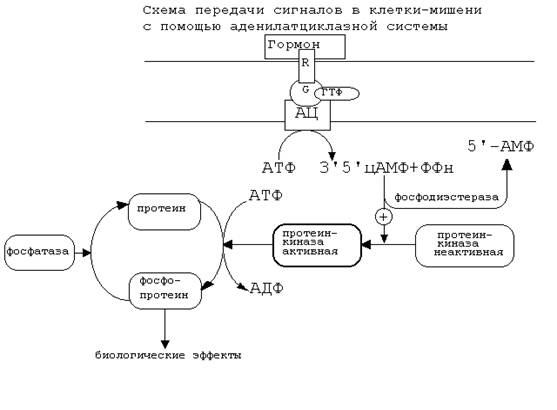
101. В зависимости от строения гормона существуют два типа взаимодействия. Если молекула гормона липофильна, (например, стероидные гормоны), то она может проникать через липидный слой наружной мембраны клеток-мишеней. Если молекула имеет большие размеры или является полярной, то ее проникновение внутрь клетки невозможно. Поэтому для липофильных гормонов рецепторы находятся внутри клеток-мишеней, а для гидрофильных - рецепторы находятся в наружной мембране. Для получения клеточного ответа на гормональный сигнал в случае гидрофильных молекул действует внутриклеточный механизм передачи сигнала. Это происходит с участием веществ, которых называют "ВТОРЫМИ ПОСРЕДНИКАМИ". Молекулы гормонов очень разнообразны по форме, а "вторые посредники" - нет. Надёжность передачи сигнала обеспечивает очень высокое сродство гормона к своему белку-рецептору. Что такое посредники, которые участвуют во внутриклеточной передаче гуморальных сигналов? Это циклические нуклеотиды (цАМФ и цГМФ), инозитолтрифосфат, кальций-связывающий белок - кальмодулин, ионы кальция, ферменты, участвующие в синтезе циклических нуклеотидов, а также протеинкиназы - ферменты фосфорилирования белков. Все эти вещества участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях. Разберем более подробно механизмы действия гормонов и внутриклеточных посредников. Существует два главных способа передачи сигнала в клетки-мишени от сигнальных молекул с мембранным механизмом действия: 1. АДЕНИЛАТЦИКЛАЗНАЯ (ИЛИ ГУАНИЛАТЦИКЛАЗНАЯ) СИСТЕМЫ 2. ФОСФОИНОЗИТИДНЫЙ МЕХАНИЗМ АДЕНИЛАТЦИКЛАЗНАЯ СИСТЕМА. Основные компоненты: мембранный белок-рецептор, G-белок, фермент аденилатциклаза, гуанозинтрифосфат, протеинкиназы. Кроме того, для нормального функционирования аденилатциклазной системы, требуется АТФ. Схема аденилатциклазной системы представлена на рисунке:
Как видно из рисунка, белок-рецептор, G-белок, рядом с которым располагаются ГТФ и фермент (аденилатциклаза) встроены в мембрану клетки. До момента действия гормона эти компоненты находятся в диссоциированнном состоянии, а после образования комплекса сигнальной молекулы с белком-рецептором происходят изменения конформации G-белка. В результате одна из субъединиц G-белка приобретает способность связываться с ГТФ. Комплекс “G-белок-ГТФ” активирует аденилатциклазу. Аденилатциклаза начинает активно превращать молекулы АТФ в ц-АМФ. ц-АМФ обладает способностью активировать особые ферменты - протеинкиназы, которые катализируют реакции фосфорилирования различных белков с участием АТФ. При этом в состав белковых молекул включаются остатки фосфорной кислоты. Главным результатом этого процесса фосфорилирования является изменение активности фосфорилированного белка. В различных типах клеток фосфорилированию в результате активации аденилат-циклазной системы подвергаются белки с разной функциональной активностью. Например, это могут быть ферменты, ядерные белки, мембранные белки. В результате реакции фосфорилирования белки могут становятся функционально активными или неактивными. Такие процессы будут приводить к изменениям скорости биохимических процессов в клетке-мишени. Активация аденилатциклазной систтемы длится очень короткое время, потому что G-белок после связывания с аденилатциклазой начинает проявлять ГТФ-азную активность. После гидролиза ГТФ G-белок восстанавливает свою конформацию и перестает активировать аденилатциклазу. В результате прекращается реакция образования цАМФ. Кроме участников аденилатциклазной системы в некоторых клетках-мишенях имеются белки-рецепторы, связанные с G-белками, которые приводят к торможению аденилатциклазы. При этом комплекс “GTP-G-белок” ингибирует аденилатциклазу. Когда останавливается образование цАМФ, реакции фосфорилирования в клетке прекращаются не сразу: пока продолжают существовать молекулы цАМФ - будет продолжаться и процесс активации протеинкиназ. Для того, чтобы прекратить действие цАМФ, в клетках существует специальный фермент - фосфодиэстераза, который катализирует реакцию гидролиза 3',5'-цикло-АМФ до АМФ. Некоторые вещества, обладающие ингибирующим действием на фосфодиэстеразу, (например, алкалоиды кофеин, теофиллин), способствуют сохранению и увеличению концентрации цикло-АМФ в клетке. Под действием этих веществ в организме продолжительность активации аденилатциклазной системы становится больше, то есть усиливается действие гормона. 102. В зависимости от строения гормона существуют два типа взаимодействия. Если молекула гормона липофильна, (например, стероидные гормоны), то она может проникать через липидный слой наружной мембраны клеток-мишеней. Если молекула имеет большие размеры или является полярной, то ее проникновение внутрь клетки невозможно. Поэтому для липофильных гормонов рецепторы находятся внутри клеток-мишеней, а для гидрофильных - рецепторы находятся в наружной мембране. Для получения клеточного ответа на гормональный сигнал в случае гидрофильных молекул действует внутриклеточный механизм передачи сигнала. Это происходит с участием веществ, которых называют "ВТОРЫМИ ПОСРЕДНИКАМИ". Молекулы гормонов очень разнообразны по форме, а "вторые посредники" - нет. Надёжность передачи сигнала обеспечивает очень высокое сродство гормона к своему белку-рецептору. Что такое посредники, которые участвуют во внутриклеточной передаче гуморальных сигналов? Это циклические нуклеотиды (цАМФ и цГМФ), инозитолтрифосфат, кальций-связывающий белок - кальмодулин, ионы кальция, ферменты, участвующие в синтезе циклических нуклеотидов, а также протеинкиназы - ферменты фосфорилирования белков. Все эти вещества участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях. Разберем более подробно механизмы действия гормонов и внутриклеточных посредников. Существует два главных способа передачи сигнала в клетки-мишени от сигнальных молекул с мембранным механизмом действия: 1. АДЕНИЛАТЦИКЛАЗНАЯ (ИЛИ ГУАНИЛАТЦИКЛАЗНАЯ) СИСТЕМЫ 2. ФОСФОИНОЗИТИДНЫЙ МЕХАНИЗМ Инозитолтрифосфат -это вещество, которое является производным сложного липида - инозитфосфатида. Оно образуется в результате действия специального фермента - фосфолипазы “С”, который активируется в результате конформационных изменений внутриклеточного домена мембранного белка-рецептора. Этот фермент гидролизует фосфоэфирную связь в молекуле фосфатидил-инозитол-4,5-бисфосфата и в результате образуются диацилглицерин и инозитолтрифосфат.
Известно, что образование диацилглицерина и инозитолтрифосфата приводит к увеличению концентрации ионизированного кальция внутри клетки. Это приводит к активации многих кальций-зависимых белков внутри клетки, в том числе активируются различные протеинкиназы. И здесь, как и при активации аденилатциклазной системы, одной из стадий передачи сигнала внутри клетки является фосфорилирование белков, которое в приводит к физиологическому ответу клетки на действие гормона. В работе фосфоинозитидного механизма передачи сигналов в клетке-мишени принимает участие специальный кальций-связывающий белок - кальмодулин. Это низкомолекулярный белок (17 кДа), на 30% состоящий из отрицательно заряженных аминокислот (Глу, Асп) и поэтому способный активно связывать Са+2. Одна молекула кальмодулина имеет 4 кальций-связывающих участка. После взаимодействия с Са+2 происходят конформационные изменения молекулы кальмодулина и комплекс “Са+2-кальмодулин” становится способным регулировать активность (аллостерически угнетать или активировать) многие ферменты - аденилатциклазу, фосфодиэстеразу, Са+2,Мg+2-АТФазу и различные протеинкиназы. В разных клетках при воздействии комплекса “Са+2-кальмодулин” на изоферменты одного и того же фермента (например, на аденилатциклазу разного типа) в одних случаях наблюдается активация, а в других - ингибирование реакции образования цАМФ. Такие различные эффекты происходят потому, что аллостерические центры изоферментов могут включать в себя различные радикалы аминокислот и их реакция на действие комплекса Са+2-кальмодулин будет отличаться. Таким образом, в роли "вторых посредников" для передачи сигналов от гормонов в клетках-мишенях могут быть: - Циклические нуклеотиды (ц-АМФ и ц-ГМФ); - Ионы Са; - Комплекс “Са-кальмодулин”; - Диацилглицерин; - Инозитолтрифосфат 106. ИНСУЛИН Основные механизмы действия инсулина: 1. Инсулин повышает проницаемость плазматических мембран для глюкозы. Этот эффект инсулина является главным лимитирующим звеном метаболизма углеводов в клетках. 2. Инсулин снимает тормозящее действие глюкокортикостероидов на гексокиназу. 3. На генетическом уровне инсулин стимулирует биосинтез ферментов метаболизма углеводов, в том числе ключевых ферментов. 4. Инсулин в клетках жировой ткани ингибирует триглицеридлипазу - ключевой фермент распада жиров. Регуляция секреции инсулина в кровь происходит с участием нейро-рефлекторных механизмов. В стенках кровеносных сосудов есть особые хеморецепторы, чувствительные к глюкозе. Повышение концентрации глюкозы в крови вызывает рефлекторную секркцию инсулина в кровь, глюкоза проникает в клетки и ее концентрация в крови снижается. Остальные гормоны вызывают повышение концентрации глюкозы в крови. Действие ИНСУЛИНА связано с повышением активности внутриклеточной фосфодиэстеразы, что приводит к снижению концентрации цАМФ и угнетению липолиза. Таким образом, инсулин усиливает синтез жира и уменьшает скорость его мобилизации.
ГЛЮКАГОН. Относится к белково-пептидным гормонам. Обладает мембранным типом взаимодействия с клеткой-мишенью. Эффект оказывает через аденилатциклазную систему. 1. Вызывает повышение активности гликоген-фосфорилазы. В результате ускоряется распад гликогена. Так как глюкагон оказывает эффект только в печени то можно сказать, что он "гонит глюкозу из печени". 2. Понижает активность гликоген-синтетазы, замедляя синтез гликогена. 3. Активирует липазу в жировых депо.
107. ГЛЮКОКОРТИКОСТЕРОИДЫ (ГКС). Относятся к стероидным гормонам, поэтому обладают внутриклеточным типом взаимодействия с клеткой-мишенью. Проникая в клетку-мишень, они взаимодействуют с клеточным рецептором и обладают следующими эффектами: 1. Ингибируют гексокиназу - таким образом они замедляют утилизацию глюкозы. В результате концентрация глюкозы в крови возрастает. 2. Данные гормоны обеспечивают процесс гликонеогенеза субстратами. 3. На генетическом уровне усиливают биосинтез ферментов катаболизма белков
108.
|
||||||||||||||
|
|
Последнее изменение этой страницы: 2016-12-28; просмотров: 657; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.43.92 (0.009 с.) |

 Клетка-мишень
Клетка-мишень
 Рецептор
Рецептор