

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Вклад генотип-средового взаимодействия в популяционную изменчивость
Очень важно понять, что два основных источника различий между людьми (генотип и среда) тесно связаны друг с другом и находятся в непрерывном взаимодействии. Различия между носителями разных генотипов могут не проявиться, если будут отсутствовать те факторы среды, которые превращают генотипические различия в видимые фенотипические. Мы уже приводили пример с различиями в цвете кожи в летний и зимний периоды. Подобных примеров можно привести множество. Так, люди с генетической склонностью к полноте вряд ли будут отличаться тучностью, если в их рационе не будет достаточного количества калорийной пищи, например, в условиях голода или при соблюдении соответствующей диеты. Люди с исключительными музыкальными способностями не смогут развить их, если они не будут иметь возможности для специальных занятий. То же самое справедливо для любой области человеческой деятельности: только в благоприятных условиях среды люди с особыми природными, наследственными задатками будут значительно отличаться от остальной популяции. Если условия среды мешают развитию задатков, мы вряд ли получим большие различия в фенотипах людей, имеющих разные генотипы. Таким образом, генотипические различия между людьми могут не превращаться в фенотипические, если среда не способствует этому. Это означает отсутствие фатальной неизбежности проявления всех генетических задатков, в том числе и неблагоприятных. Известно, например, что такое заболевание, как шизофрения, имеет значительный наследственный компонент, однако носители неблагоприятных генов могут избежать заболевания, если не будут подвергаться значительным стрессогенным воздействиям среды, которые чаще всего и "запускают" болезнь.
Итак, имеющиеся между людьми различия в генотипах могут проявляться или не проявляться в зависимости от средовых условий - в одном средовом диапазоне эти различия будут явно выражены, тогда как в другом мы и не заподозрим, что они существуют. Если перевести эти рассуждения на язык дисперсий, мы можем сказать так: при одном и том же составе генотипов могут существовать условия среды, которые способны менять дисперсию фенотипов в популяции. Если условия среды приводят к увеличению фенотипических различий между носителями разных генотипов (например, доступность калорийной пищи в примере со склонностью к полноте), результатом будет увеличение фенотипической дисперсии в популяции. Если же среда нивелирует различия между индивидами, фенотипическая дисперсия будет уменьшаться.
Итак, взаимодействие конкретных генотипов со средой может приводить к изменению фенотипической дисперсии. Этот феномен носит название генотип-средового взаимодействия. Поскольку генотип-средовое взаимодействие способно влиять на дисперсию признака в популяции, из этого следует, что оно порождает определенную долю фенотипической дисперсии.
Проблема вклада генотип-средового взаимодействия в количественную изменчивость начала обсуждаться достаточно давно. Дж. Холдейн (Haldane J., 1946; цит. по: Эрман Л. и Парсонс П., 1984) рассматривает такой пример. Допустим, в популяции существуют два генотипа - А и В. У этих генотипов в условиях среды X и Y проводят измерения какого-то количественного признака. Полученные количественные значения в зависимости от степени выраженности признака обозначают цифрами от 1 до 4. Число сочетаний факторов А, В, X и Y равно числу возможных способов перестановки, т.е. и в данном случае равняется 24 (4! = 1х2х3х4 = 24). Однако если условиться, что генотип А в среде X всегда имеет большие значения признака, чем генотип В, то остается только шесть возможных комбинаций, для которых Дж.Холдейн приводит соответствующую таблицу. Мы же попробуем изобразить все шесть ситуаций графически (рис. 4.10).
Рассмотрим первый и второй графики. Мы видим, что и в первой, и во второй средах генотип А имеет более высокие значения признака, чем генотип В, причем прямые параллельны, а это означает, что дисперсия, т.е. разброс значений фенотипов, во всех средах остается одинаковой. Таким образом, генотип-средовое взаимодействие как компонента дисперсии отсутствует. Теперь посмотрим, имеется ли реальное взаимодействие генотипа и среды в соответствии с представлениями о норме реакции. Мы видим, что при переходе из среды X в среду Y для каждого из генотипов произошло изменение количественных значений признака. При этом генотипы отреагировали на изменение среды сходным образом, сохранив свои относительные положения. На этом примере ясно видно, что взаимодействие генотипа и среды может пониматься двояко. С одной стороны, существует реальное взаимодействие каждого генотипа со средой, приводящее к изменению конкретного фенотипа. С другой стороны, имеется эффект статистического взаимодействия факторов наследственности и среды, влияющий на величину соответствующего компонента дисперсии. В данном случае, несмотря на существование реального взаимодействия конкретных генотипов и конкретных условий среды, компонента фенотипической дисперсии, обусловленная взаимодействием двух факторов, оказалась равной нулю. Этот момент необходимо четко уяснить, поскольку очень часто реальное взаимодействие генотипа и среды смешивается со статистическим эффектом взаимодействия факторов.
Первый и второй график и сходны, различаются лишь направления изменения фенотипов при переходе из среды X в среду Y. Для первого варианта примером может служить такая реальная ситуация: ученики младших классов, обучавшиеся у молодого неопытного учителя, попадают к сильному педагогу, и при этом все улучшают свои показатели, но сохраняют те же ранговые места. Обратная ситуация может быть примером для второго графика. Можно представить себе и такую ситуацию. Люди с высокой и низкой тревожностью будут сходно реагировать на ситуацию неопределенности и тревоги ухудшением показателей деятельности, но деятельность первых будет страдать сильнее, чем деятельность вторых.
Третий график отличается от первых двух тем, что линии здесь пересекаются. Соответственно по своим фенотипическим значениям в среде X генотип А будет опережать В, а в среде Y - наоборот. Оба генотипа при переходе из среды X в среду Y будут давать прирост фенотипических значений признака, но в разной мере. Генотип В оказывается более чувствительным к среде, соответственно он дает и больший прирост, сразу на четыре единицы, тогда как генотип А - всего на две. В этом случае генотип-средовое взаимодействие как компонента дисперсии будет отличным от нуля. Примером такого рода взаимодействия может послужить такая ситуация, когда общительный (А) и необщительный (В) ребенок во время болезни находятся дома (среда У) или в больнице (среда X). Дети более комфортно чувствуют себя в домашних условиях, в то же время общительного ребенка вынужденная изоляция от сверстников тяготит, однако в больнице он будет лучше адаптироваться, чем необщительный ребенок.
На четвертом графике генотип А в обоих средах имеет большие фенотипические значения признака, но генотипы по-разному реагируют на переход из среды X в среду Y. Для генотипа А характерно уменьшение значений, а для генотипа В - увеличение. Здесь также присутствует генотип-средовое взаимодействие как компонент дисперсии. Воспользуемся примером, который приводит Дж. Холдейн. Он рассматривает нормальных (А) и умственно отсталых (В) детей, которые обучаются в обычной (среда X) и специализированной (среда Y) школах. Понятно, что в обычной школе умственно отсталые дети будут значительно отставать от обычных детей, тогда как в специализированной школе с программой, рассчитанной на умственно отсталых детей, последние дадут прирост успешности, а нормальные дети будут отставать от своих сверстников.
На пятом графике опять наблюдается пересечение линий. При переходе из среды X в среду Y генотип А демонстрирует уменьшение значений, тогда как генотип В - увеличение. В среде X фенотипические различия между генотипами больше, чем в среде Y. Опять мы видим, что генотип-средовое взаимодействие меняет дисперсию фенотипов. В качестве примера можно рассмотреть людей с высоким (А) и низким (В) уровнями активации. В условиях тишины и изоляции (X) более эффективно будут работать первые, тогда как в условиях шума (Y) - вторые.
И последний, шестой вариант. Линии на графике опять пересекаются. Генотип В более чувствителен к изменению среды и демонстрирует рост величины признака при переходе в среду Y, тогда как генотип А менее чувствителен к среде и реагирует иначе. Генотип-средовое взаимодействие вносит вклад в дисперсию признака. Возможным примером может служить ситуация, в которой генотипам А и В соответствуют дети с разными типами способностей. Одни дети (генотип А) обладают средним уровнем способностей, при опережающем развитии пространственных способностей, а другие (генотип В) имеют ярко выраженные вербальные способности и испытывают серьезные затруднения при решении пространственных задач. В соответствии с этим дети будут демонстрировать разную успешность на уроках геометрии (X) и литературы (Y). Все эти примеры наглядно демонстрируют, что изменчивость в популяции складывается не только за счет простого суммирования эффектов среды и генотипа, но и включает компонент генотип-средового взаимодействия. Наличие генотип-средового взаимодействия увеличивает популяционную дисперсию признака. Это связано с тем, что представители различных генотипов по-разному реагируют на одни и те же изменения среды. Это и приводит к увеличению популяционной изменчивости.
Чтобы обнаружить статистический компонент Г-С взаимодействия необходимо подобрать группы индивидов, отличающихся своими генетическими конституциями, и находящихся в различных типах условий среды. В психогенетических исследованиях, как правило, так и поступают. Например, при исследованиях аффективных расстройств (Kendler K.S et al., 1995, цит. по Равич-Щербо И.В. и др., 1999) можно подобрать группы испытуемых из генетически отягощенных и неотягощенных семей. Это и будут контрастные генотипы ("плохой" и "хороший"). Внутри каждой из этих групп можно подобрать индивидов, находящихся в контрастных средах (благоприятной и неблагоприятной). Таким образом, мы будем иметь 4 группы: в первой из них "хороший" генотип будет сочетаться с "хорошей" средой, во второй - "хороший" генотип будет сочетаться с "плохой" средой, в третьей - "плохой" генотип будет находиться в "хорошей" среде и, наконец, в четвертой - и генотип, и среда будут "плохими" (табл. 4.3).
Табл. 4.3.
Схема эксперимента
|
| Неотягощенный генотип (G+)
| Отягощенный генотип (G-)
| | Благоприятная среда (Е+)
| G+ E+
| G- E+
| | Неблагоприят-ная среда (Е-)
| G+ E-
| G- E-
| Это классический пример планирования эксперимента для модели двухфакторного дисперсионного анализа с целью выявления эффекта взаимодействия факторов. Если в таком эксперименте мы выявим разную чувствительность контрастных генотипов к средовым изменениям, то можно делать заключение о наличии Г-С взаимодействия (рис. 4.11).

Г-С взаимодействие легко обнаруживается в экспериментах на животных. Классическим примером является разная обучаемость в лабиринте селектированных "умных" и "глупых" крыс в зависимости от условий выращивания молодняка (рис. 4.12)

Механизм увеличения фенотипической дисперсии при наличии Г-С взаимодействия можно наглядно продемонстрировать на примере депрессивного расстройства, которое можно отнести к разряду психических заболеваний с наследственной предрасположенностью и пороговым эффектом. В популяции существуют индивиды с различной наследственной предрасположенностью к заболеванию (рис 4. 13).

Часть из них находится в острой стадии болезни, т.е. за условным порогом (правая часть распределения за чертой). Предрасположенные из группы риска имеют гораздо больше шансов оказаться за порогом болезни, т.е. попасть в группу больных. Вместе с тем имеются индивиды, абсолютно не подверженные депрессии - они располагаются в противоположном конце распределения (слева). Допустим, вся популяция начинает подвергаться неблагоприятным условиям среды (E - обозначено стрелкой внизу слева), способствующим развитию депрессии (например, увеличивается стрессогенность среды). Тогда можно предположить, что индивиды, устойчивые к развитию депрессии, совсем не изменят свой фенотип, т.е. не будут проявлять даже малейших признаков депрессии. Это означает, что левая часть распределения не будет смещаться в сторону болезни. Что же будет происходить с остальными представителями популяции? Можно ожидать, что чувствительность к стрессогенной среде будет нарастать по мере приближения к порогу, и индивиды будут проявлять тем более признаков депрессии в своем фенотипе, чем ближе они к группе риска. Смещение фенотипов на графике показано стрелками разной длины. Чем более чувствителен генотип, тем длиннее стрелка. Таким образом, из рисунка становится понятно, что все распределение при переходе популяции в неблагоприятную среду начинает как бы растягиваться, причем левый его край стоит на месте, а правый - сдвигается в сторону болезни. Пунктирная линия на графике представляет собой новое распределение фенотипов в измененной среде: оно более растянуто, что говорит об увеличении фенотипической дисперсии; кроме того, заметно, что гораздо больше индивидов окажется в зоне болезни (за пороговой чертой). Этот пример демонстрирует, что при разной реакции генотипов на изменение среды фенотипическая дисперсия меняется именно за счет компонента Г-С взаимодействия.
В предыдущих темах мы рассматривали упрощенные модели, в которых принималось, что влияния генотипа и среды аддитивны. В таких случаях фенотипическая дисперсия представляет собой сумму генетической и средовой компоненты (VP = VG + VE). Если же имеют место Г-С взаимодействия, то в формулу фенотипической структуры популяции должен быть добавлен еще один компонент (VGE - дисперсия, обусловленная генотип-средовым взаимодействием). Тогда она примет следующий вид:
VP = VG + VE + VGE.
О наличии Г-С взаимодействия свидетельствуют и различия в значениях коэффициентов наследуемости в контрастных средах. Например, при изучении наследуемости алкоголизма (Heath A.S. et al., 1989, цит. по Равич-Щербо И.В. и др., 1999) было обнаружено, что в популяции замужних женщин он оказывается ниже, чем в популяции незамужних. Более высокий коэффициент наследуемости говорит о более ярком проявлении генетической вариативности в среде, соответствующей ситуации незамужней женщины. Следовательно, в ситуации замужества наследственные различия по предрасположенности к алкоголизму, по-видимому, не проявляются столь заметно в фенотипических различиях. Из этого можно заключить, что отягощенный генотип должен быть более чувствителен к изменениям среды (рис. 4.14).

Еще раз подчеркнем, что следует различать генотип-средовое взаимодействие в статистическом смысле и реальное взаимодействие генотипа и среды при формировании конкретного фенотипа.
Генотип-средовая ковариация
Рассмотренное нами генотип-средовое взаимодействие следует отличать от другого явления, с которым его часто путают. Речь идет о генотип-средовой ковариации (или корреляции). Генотип-средовая ковариация имеет место тогда, когда генотипы оказываются в определенных средах не случайно, а в соответствии со своей наследственной предрасположенностью. Это означает, что генотипы неравномерно распределяются по разным средам. Примером может служить соответствие различных видов, рас, подвидов, разновидностей различным местам обитания. Приспособленность генотипов к определенным условиям среды приводит к тому, что генотипы концентрируются в тех средах, которые больше соответствуют их особенностям. В человеческих популяциях наблюдаются похожие явления. В таких различных средах, как стадион, концертный зал или библиотека, мы, скорее всего, встретим людей с определенными генетическими предрасположенностями: на стадионе будут чаще встречаться люди с природной склонностью к двигательной деятельности, в концертном зале - люди с музыкальной одаренностью, а в библиотеке будет больше людей с высокими умственными способностями.
Мы видели, что генотип-средовое взаимодействие всегда увеличивает популяционную дисперсию. Генотип-средовая ковариация может как увеличивать, так и уменьшать дисперсию фенотипов в популяции. В жизни человека генотип-средовые ковариации играют важную роль. Родители могут передавать ребенку не только определенную генетическую предрасположенность, но и одновременно обеспечивать ему соответствующую среду для развития природных задатков. Здесь мы имеем дело с культурной преемственностью, которую не всегда легко отличить от наследственности. Например, родители-музыканты большей частью заботятся о том, чтобы их дети также получили музыкальное образование. В таких семьях можно ожидать, что дети вместе с "генами музыкальности" унаследуют от родителей и "музыкальную среду".
Генотип-средовая ковариация, как и генотип-средовое взаимодействие, влияет на общую дисперсию признака, а следовательно, может влиять на величину оценок наследуемости. Это следует учитывать при проведении исследований в генетике поведения.
Рассмотрим явление генотип-средовой ковариации более подробно. Сначала обратимся к математике, чтобы понять, что генотип-средовая ковариация является компонентом дисперсии фенотипов.
Мы знаем, что фенотип представляет собой результат совместного действия генотипа и среды. Если перевести это на язык статистики, то количественное значение признака у конкретного i -того индивида можно представить как фенотипическое отклонение (Pi) от популяционной средней. Такое значение фенотипа есть результат, с одной стороны, генотипического отклонения (Gi) от популяционной средней и, с другой стороны, - средового отклонения (Ei). Таким образом, можно записать языком математики, что конкретный фенотип есть результат взаимодействия генотипа и среды:
Pi = Gi + Ei.
Чтобы определить дисперсии отклонений, нужно каждое i -е отклонение возвести в квадрат, просуммировать все квадраты отклонений и суммы разделить на n.

Величина  представляет собой общую фенотипическую дисперсию, представляет собой общую фенотипическую дисперсию,  - генетическая дисперсия, - генетическая дисперсия, 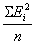 - средовая дисперсия. Поскольку Gi и Ei представляют собой генотипические и средовые отклонения, то сумма их произведений - средовая дисперсия. Поскольку Gi и Ei представляют собой генотипические и средовые отклонения, то сумма их произведений 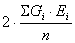 есть ковариация генотипа и среды. Итак, есть ковариация генотипа и среды. Итак,
VP = VG + VE + 2covGE.
Если считать, что генотипы равномерно распределены по средам (т.е. оказываются в определенной среде совершенно случайно), то компонент 2covGE становится равным 0, и тогда мы опять имеем ту формулу фенотипической структуры популяции, которая рассматривалась нами как исходная:
VP = VG + VE.
В реальных популяциях генотип-средовая ковариация является обычным явлением и ее вклад в вариативность признаков, как правило, не равен нулю. Действительно, трудно себе представить, чтобы индивид совершенно случайно попадал в ту или иную среду. С рождением каждого ребенка популяция пополняется еще одним генотипом, но ребенок обычно сразу же попадает в среду своей собственной семьи, к своим собственным родителям, бабушкам, дедушкам, братьям и сестрам, с которыми у него есть общие гены. Если при игре в рулетку брошенный шарик случайно оказывается в ячейке, то только что родившийся ребенок занимает "ячейку" вполне определенную: в своей семье, своем городе, своей стране. Таким образом, его среда не может быть совершенно случайной.
Рассмотрим следующую схему (рис. 4.15).
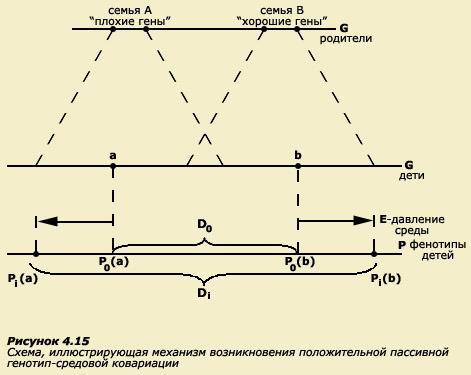
Предположим, в популяции существуют две семьи (А и В). Супруги А, оба, обладают "плохими" генами в отношении какого-то количественного признака с аддитивным типом наследования (допустим, это интеллект), т.е. у них преобладают гены-ослабители. В то же время в другой семье (В) супруги имеют "хорошие" гены (преобладают гены-усилители). В результате рекомбинаций при образовании гамет дети супругов А и супругов В могут получить различные сочетания генов, и, в принципе, среди них могут оказаться обладатели сходных сочетаний (зона перекрытия на рисунке), однако все же вероятность того, что дети супругов А окажутся обладателями "худших" генов достаточно велика, и, наоборот, дети супругов В скорее всего получат "лучшие" гены. На схеме генотипы детей обозначены точками а и b на соответствующей оси (внизу). Теперь представим себе, в какой среде окажутся дети в этих семьях. Скорее всего в силу более высоких умственных способностей родители В обеспечат своему ребенку лучшую среду для развития, чем родители А. В семье В среда будет обогащенной (больше книг, развивающих игрушек, дополнительных занятий, речевое общение более высокого уровня и т.д.), а в семье А - обедненной, поскольку у родителей, вероятно, будет меньше возможностей создать подобную среду.
Следовательно, среда семьи В будет способствовать максимальному развитию исходных задатков своего ребенка, и его фенотип будет постоянно улучшаться. В то же время среда семьи А не создаст возможностей для развития фенотипа ребенка. В результате наследственно заложенные задатки не проявятся в полной степени. Таким образом, мы имеем ситуацию, в которой совершенно пассивно, просто по факту своего рождения, обладатель лучшего генотипа унаследовал и лучшую среду, а обладатель худшего генотипа унаследовал худшую среду.
Здесь мы имеем дело с явлением, которое в психогенетике называется пассивной положительной ковариацией генотипа и среды. Ковариация (корреляция) всегда положительна, если два параметра изменяются (ковариируют) в одном направлении. Например, чем больше рост, тем больше длина ступни человека. Это пример положительной ковариации. В рассмотренном нами случае генотип и среда также ковариируют положительно, поскольку лучшему генотипу соответствует лучшая среда и наоборот. Пассивной ковариация генотипа и среды называется потому, что ни родители, ни дети не прикладывают специальных усилий для создания среды: дети просто "наследуют" среду так же, как и гены. Это называется культурным наследованием.
На той же схеме (рис. 4.15) стрелками показано, что среда как бы "тянет" фенотипы (Pa) и (Pb) в разные стороны. Понятно, что в результате та разница, которая имелась между детьми при рождении (диапазон фенотипических различий - D0), будет усугубляться (диапазон фенотипических различий - Di). Если такая картина наблюдается не только в одной семье, а типична для популяции вообще, то результатом генотип-средовой ковариации будет увеличение фенотипической дисперсии, поскольку разброс между фенотипами в результате такой ковариации, как мы видим, возрастает.
Теперь рассмотрим еще одну подобную схему (рис. 4.16).

Предположим, что в семье В родился второй ребенок (b2), которому "не повезло", и он получил худшее сочетание генов. Родители обнаружили, что второй ребенок не столь одарен от природы, как первый, и направили все усилия на его развитие, забросив первого, который "и сам пробьется". Тем самым они создали худшему генотипу лучшие условия среды. Это пример отрицательной генотип-средовой ковариации, поскольку переменные варьируют в различных направлениях (хуже генотип - лучше среда и, наоборот, лучше генотип - хуже среда). В результате фенотипические различия между детьми начнут уменьшаться (конечный диапазон различий Di меньше начального - D0). Таким образом, отрицательная генотип-средовая ковариация должна уменьшать популяционную дисперсию, но это в том случае, если рассмотренный пример представляет собой типичное явление. Напоминаем, что ковариация является величиной статистической и может быть обнаружена только на значительных выборках. Если же подобные усилия родителей наблюдаются лишь в одной семье, а в других происходит нечто иное, то никакой отрицательной ковариации генотипа и среды мы не получим.
Только что рассмотренная ситуация является примером отрицательной реактивной ковариации генотипа и среды. Реактивной обычно называется ковариация, возникающая в результате специальных усилий социального окружения ребенка в ответ на какие-либо его специфические особенности, связанные с наследственной конституцией. Например, дети, проявляющие признаки природной одаренности в спорте, музыке, изобразительной деятельности, привлекают внимание взрослых (родителей, педагогов, воспитателей), и те стараются создать условия для дальнейшего развития природных задатков: помещают детей в специализированные школы, кружки по интересам и т.п. В результате "хороший" генотип получает "хорошую" среду, т.е. наблюдается положительная ковариация.
Реактивная ковариация может быть и отрицательной. Она может возникать, например, в том случае, когда наследственной отягощенности или какому-либо неблагополучию ребенка, обусловленному наследственными причинами, противопоставляются компенсирующие усилия родителей, врачей, педагогов, заставляющие работать среду против генотипа. Однако это скорее исключение, чем правило. Большинство детей с проблемами развития оказываются в среде, которая только усугубляет их неблагополучие. Например, большие трудности часто возникают у детей с гиперактивностью и нарушениями внимания, нарушениями эмоционально-волевой сферы, "трудным" темпераментом. Все это можно назвать синдромом "трудного" ребенка. К сожалению, такие "особые" дети чаще вызывают отрицательные реакции окружающих: их больше наказывают, унижают и обижают, они реже бывают успешными в различных видах деятельности. Получается, что их природное (чаще всего оно и наследственное) неблагополучие влечет за собой и неблагоприятную среду. Ковариация генотипа и среды в этом случае будет положительной, поскольку и генотип, и среда действуют в одном направлении ("плохой" генотип влечет за собой "плохую" среду). Безусловно, это приводит к отрицательным последствиям для фенотипа, но генотип-средовая ковариация в данном случае положительная.
Усилия практических психологов должны быть направлены на преодоление существующей тенденции. Необходимо помогать родителям, воспитателям и педагогам создавать такие условия среды, которые будут улучшать фенотип ребенка, имеющего ту или иную отягощенность. Практический психолог, участвуя в отборе детей в специализированные школы, консультируя родителей, участвуя в организации среды в детских учреждениях, должен помнить, что от его усилий зависит, произойдут ли положительные сдвиги в развитии ребенка. Поэтому при "плохом" генотипе следует создавать отрицательную ковариацию генотипа и среды, а при "хорошем" - положительную.
Мы рассмотрели два типа ковариации - пассивную и реактивную - и обнаружили, что она может быть как положительной, так и отрицательной. Если вернуться к формуле:
VP = VG + VE +2covGE,
то становится понятным, что положительная ковариация должна увеличивать фенотипическую дисперсию в популяции, а отрицательная - уменьшать.
В психогенетике, помимо пассивной и реактивной форм ковариации генотипа и среды, выделяют еще и активную ковариацию. Она характерна для более старших возрастов, начиная с подросткового, когда индивид самостоятельно (активно) выбирает ту среду, которая ему более подходит. Часто это происходит против воли родителей и других взрослых. Типичным примером активной генотип-средовой ковариации является судьба самого Ф. Гальтона, который по настоянию отца долгое время готовился к карьере врача, но после смерти отца забросил медицину и хотя и продолжал изучать природу человека, но уже в совершенно ином аспекте. Таким образом, получается, что не столько среда влияет на формирование индивидуальности человека, сколько сама индивидуальность (с ее уникальной генетической конституцией) формирует свою особую среду. Впервые на это обстоятельство обратила внимание американская исследовательница Сандра Скарр в 1983 г. (Scarr S., McCartney K., 1983).
Наличие генотип-средовой ковариации приводит к тому, что средовые условия у более близких родственников, имеющих высокую долю общих генов (т.е. сходные генотипы), оказываются более сходными, чем у более отдаленных родственников. Характерным примером могут служить идентичные близнецы, разлученные в раннем детстве и никогда не встречавшиеся друг с другом. Уже будучи солидными людьми, при первой встрече они с удивлением обнаруживают множество совпадающих обстоятельств своей жизни: сходство профессий, привычек, любимых занятий и даже сходные особенности спутников жизни.
Можно ли оценить вклад генотип-средовой ковариации в изменчивость признака? В психогенетике для этого существуют специальные методы. Большинство из них используют приемы, основанные на сравнении оценок характеристик среды у родственников различной степени родства. Одним из наиболее распространенных является методика интервью и наблюдений НОМЕ (Home Observation of Measurement of the Environment), позволяющая оценить эмоциональные и вербальные реакции родителей, отношение к наказаниям, организацию физической среды в семье и т.п. Используются также всевозможные опросники, в которых родственники оценивают различные параметры семейной и внесемейной среды. Полученные оценки могут сравниваться у родственников подобно тому, как это делается для любых количественных характеристик фенотипа. Так можно оценить сходство родственников не только по фенотипу, но и по средовым параметрам. Если средовое сходство оказывается выше у более близких родственников, можно предполагать наличие генотип-средовой ковариации.
(В качестве дополнительного материала см. хрестоматии 4.1. и 4.2.) (см. Видео)
Выводы
- 4.1. Количественная изменчивость и методы ее описания
- Требования к психологическим измерениям в генетике поведения соответствуют основным требованиям психометрики (надежность, валидность, репрезентативность).
- Распределение частот встречаемости различных количественных значений признака в популяции характеризуются двумя статистическими величинами - центральной тенденцией (мода, медиана, среднее) и разбросом значений вокруг среднего (дисперсия).
- Дисперсия характеризует межиндивидуальные различия (изменчивость, вариативность).
- Генетика поведения изучает природу индивидуальных различий.
- 4.2. Наследственность и среда как факторы возникновения количественной изменчивости
- Вся совокупность генов организма составляет его генотип.
- Любые проявления организма в каждый момент жизни составляют его фенотип.
- Фенотип есть результат взаимодействия генотипа со средой.
- Количественная изменчивость может возникать в результате полимерного действия многих генов на один признак.
- Дисперсия количественного признака, возникающая за счет действия генов, носит название генетической дисперсии.
- Существуют различные формы взаимодействия генов: аддитивное, доминирование, эпистаз и др.
- Количественная изменчивость может возникать под действием факторов среды.
- Дисперсия количественного признака, возникающая за счет средовых влияний, носит название средовой дисперсии.
- Генотипы по-разному реагируют на одни и те же изменения среды. Специфический характер реакции данного гeнотипа на изменение окружающих условий носит название нормы реакции.
- Генотипы отличаются по своей чувствительности к среде.
- Фенотипическая изменчивость в популяции складывается из генетической и средовой изменчивости; фенотипическая популяционная дисперсия признака представляет собой сумму его генетической и средовой дисперсий.
- 4.3. Показатель наследуемости и его особенности
- Доля генетической составляющей в фенотипической дисперсии признака называется наследуемостью.
- Наследуемость не является атрибутом признака как такового, а зависит от состава генотипов в популяции и от конкретных средовых условий.
- Высокая наследуемость не означает невозможности изменения признака при изменении среды.
- Наследуемость есть характеристика популяции, а не конкретного индивида и его конкретного фенотипа.
- 4.4. Генотип-средовое взаимодействие
- Существующие в популяции генетические различия могут не превращаться в фенотипические, если среда не способствует этому.
- Генотип-средовое статистическое взаимодействие увеличивает фенотипическую дисперсию в популяции.
- Генотип-средовое взаимодействие является компонентом фенотипической дисперсии.
- Следует различать генотип-средовое взаимодействие как статистический компонент дисперсии и реальное взаимодействие генотипа и среды при формированиии конкретного фенотипа.
- Явление неслучайного распределения генотипов по средам носит название генотип-средовой ковариации (корреляции).
- Генотип-средовая ковариация может быть положительной, если и генотип и среда варьируют в одном направлении (чем хуже генотип, тем хуже среда; чем лучше генотип, тем лучше среда) и отрицательной, если генотип и среда варьируют в противоположных направлениях (чем хуже генотип, тем лучше среда и наоборот).
- Положительная ковариация увеличивает популяционную дисперсию, отрицательная - уменьшает.
- Различают три вида генотип-средовой ковариации: пассивную, реактивную и активную.
- Генотип-средовое взаимодействие и генотип-средовая ковариация могут влиять на величины оценок наследуемости.
Словарь терминов
- Дисперсия
- Мода
- Медиана
- Стандартное отклонение
- Решетка Пеннета
- Эпистаз
- Локус
- Аллель
- Полимерные гены
- Наследуемость
- Норма реакции
- Инбридинг
- Клон
- Генотип-средовое взаимодействие
- Генотип-средовая ковариация
Вопросы для самопроверки
- Какие статистические величины существуют для описания частотных распределений?
- Почему тесты, применяемые для измерений в психогенетике, должны отличаться надежностью и валидностью?
- Что такое дисперсия и по какой формуле она вычисляется?
- Какие статистические величины используются для характеристики групповых и индивидуальных различий?
- Может ли возникать нормальное распределение признака в популяции при отсутствии генетической изменчивости?
- В каких группах может отсутствовать генетическая изменчивость?
- Какие группы организмов называются клонами?
- Существуют ли клоны в человеческой популяции?
- Что в генетике называется нормой реакции?
- Почему нежелательно в определении нормы реакции пользоваться такими понятиями как предел, предельные возможности генотипа и т.п.?
- Приведите примеры нормы реакции у животных и растений.
- Почему говорят, что наследуется не признак, а норма реакции?
- Почему невозможно получить нормы реакции для человека?
- Графики каких гипотетических норм реакции для психологических признаков человека можно построить?
- Что является критическим для нормального развития интеллекта у человека?
- Почему психогенетика работает в основном с дисперсиями?
- От каких факторов зависит количественная изменчивость признаков?
- Что такое генотип и фенотип? Приведите примеры по
|




