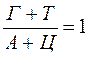Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Уровни изучения неслественностиСодержание книги
Поиск на нашем сайте
Главным и единственным методом изучения наследственности организмов является классический генетический (гибридологический) анализ, или, как его еще называют, формальный генетический анализ. Основы этого метода были разработаны Г. Менделем. Этот метод заключается в последовательном разложении генома анализируемого организма на группы сцепленных генов, а групп сцепления — на генные локусы с дальнейшим установлением последовательности генных локусов вдоль хромосомных пар и выяснением тонкой структуры генов. Генетический анализ в принципе подобен химическому анализу, задача которого заключается в разложении сложных химических соединений на более простые компоненты. Однако в отличие от химического анализа, например нуклеопротеидов, расщепление которых на структурные части основано на гидролизе, классический генетический анализ основывается на расщеплении (сегрегации) и рекомбинации генов в мейозе и осуществляется путем скрещиваний особей с разными признаками и учета результатов скрещиваний. Схема генетического анализа организмов состоит из ряда последовательных этапов, а именно: 1. Идентификация генов. 2. Установление генных локусов на хромосомных парах. 3. Установление последовательности генных локусов вдоль хромосомных пар. 4. Выяснение тонкой структуры генов. Результаты генетического анализа оформляют путем составления генетических карт. Одним из важнейших показателей эффективности генетического анализа является его разрешающая способность, которая в общих чертах может быть аналогизирована с разрешающей способностью оптических методов исследования. Подобно тому, как разрешающая способность оптических приборов (микроскопов) ограничена волновой природой света, разрешающая способность генетического анализа ограничивается количеством исследуемого потомства, получаемого в скрещиваниях, ибо чем большим является количество потомства, тем большей является возможность обнаружения среди них редких рекомбинантов и, следовательно, установления частоты кроссинговера. Начиная с 1910 г., в генетике в качестве экспериментальной модели (системы) широко используют плодовую мушку Drosophila melanogaster (рис. 103). Являясь эукариотом с дифференцированными тканями, этот организм очень удобен для изучения многих вопросов наследственности. В частности, у этого организма было идентифицировано и изучено большое количество генных и хромосомных мутаций, причем хромосомные мутации из-за больших размеров в клетках слюнной железы оказались доступными для изучения с помощью обычного микроскопа. На этом организме была показана «мощь» генетического анализа. Однако разрешающая способность генетического анализа всегда имеет ограничения, поскольку возможность получения большого количества потомства всегда ограничена до определенных пределов даже у тех видов, у которых оно составляет сотни организмов на пару, как, например, у D. melanogaster. Поэтому у организмов, размножающихся половым путем, в том числе и у плодовой мушки возможно выполнение лишь трех первых этапов генетического анализа.
В качестве экспериментальных моделей широко используют также дрожжи. Являясь простейшими эукариотами, эти организмы обладают всеми преимуществами бактерий. Но кроме этого, они оказались доступными для изучения на них генетики митохондрий, сплайсинга РНК, гаплои-дии и диплоидии.
Длительное время для изучения генетического контроля развития животных организмов использовали D. melanogaster. Однако, начиная с 60-х гг., в качестве модельного объекта в генетике развития стали использовать круглого гельминта Caenorhabditis (рис. 104). Имея длину в 1 мм, эта нематода состоит примерно из 1000 клеток. Ее генетический аппарат представлен 6 парами гомологичных хромосом, на которых локализовано около 3000 генов. В гаплоидном состоянии геном состоит из 8´107 пар нуклеотидов. Что касается растений, то для изучения генетики развития этих организмов используют травянистое растение Arabidopsis thaliana (рис. 105). Преимущества этого растения в качестве экспериментальной модели заключаются в том, что его легко культивировать в лабораторных условиях и что оно имеет очень короткий срок вегетации (всего лишь 5 недель). Кроме того, геном этого растения состоит из 7´107 нуклеотидных пар. У всех этих организмов идентифицированы различные мутации, созданы их геномные библиотеки и секвенировано большинство генов. Секвенирование стало методом изучения тонкого строения генов у всех организмов. Развитие молекулярной биологии привело к разработке методологии генетической инженерии, которая нашла исключительно широкое применение в животноводстве, растениеводстве, а также в изучении нормальной и патологической наследственности человека (см. раздел V).
Вопросы для обсуждения
1. Дайте определение наследственности и объясните, каким образом наследственность определяет непрерывность жизни? 2. Является ли изменчивость свойством живого и если да, то почему? 3. Какие формы изменчивости вы знаете? 4. Что важнее, наследственность или среда? 5. Дайте определение генотипа и фенотипа. 6. В чем заключается классический генетический анализ и применим ли он для изучения наследственности всех организмов? Каковы его возможности и ограничения? 7. Для чего используют в изучении наследственности и изменчивости экспериментальные модели? 8. Какие организмы используются в генетике в качестве моделей для изучения генетических закономерностей? 9. На каких уровнях изучают наследственность и изменчивость? 10. Каково значение молекулярно-генетических исследований наследственности и изменчивости? 11. Можно ли изучать тонкое строение генов, не прибегая к скрещиваниям?
Литература
Дубинин Н. П. Генетика. Кишинев: Штиинца. 1986. 534 стр. Грин Н., Стаут У., Тейлор Д. Биология. М.: Мир. 1996. 386 стр. Schleif R. Genetics and Molecular Biology. The Johns Hopkins University Press. 1993. 698 pp.
Глава X ГЕНЕТИЧЕСКИЙ МАТЕРИАЛ
В соответствии с современными представлениями генетическим материалом являются нуклеиновые кислоты. Нуклеиновые кислоты были обнаружены в ядрах клеток в 1869 г. швейцарским физиологом Фридрихом Мишером. Это открытие является настолько важным, что оно заслуживает приведения здесь цитаты из работы Ф. Мишера, в которой он описывал свои опыты, а именно: «Обрабатывая клетки гноя слабыми щелочными растворами, я получил в результате нейтрализации раствора осадок, который не растворялся ни в воде, ни в уксусной кислоте, ни в разведенной соляной кислоте, ни в обычном солевом растворе и который не мог принадлежать ни к одному из белков, известных в настоящее время». Обнаруженное вещество Ф. Мишер назвал «нуклеином». Как считают, он не мог знать, что открыл ДНК и что оказался в начале исследований ДНК. Но, определяя заслуги Ф. Мишера в качестве первооткрывателя нуклеиновых кислот, нельзя не отметить, что первое предположение о роли нуклеиновых кислот в качестве генетического материала было сформулировано в 1914 г. доцентом Петербургского университета А. Щепотьевым. Материалом, из которого состоят гены, является дезоксирибо-нуклеиновая кислота (ДНК), а материалом, обеспечивающим декодирование генетической информации, являются РНК. У отдельных вирусов первичным генетическим материалом является рибонуклеиновая кислота (РНК). Таким образом, нуклеиновые кислоты являются хранителями (носителями) и переносчиками генетической информации. Сложившиеся представления о том, что первичным генетическим материалом у абсолютного большинства живых существ является ДНК, основаны на ряде прямых и косвенных доказательств, среди которых исторически самым первым прямым доказательством генетической роли ДНК является установленная еще в 1944 г. способность ДНК трансформировать пневмококки из одного типа в другой. К настоящему времени трансформация установлена у микроорганизмов многих видов. В 50-е годы прямые доказательства генетической специфичности ДНК были получены также в результате изучения размножения бактериальных вирусов в бактериях и переноса ими генетической информации с помощью ДНК от одних бактериальных клеток к другим. Это явление известно под названием трансдукции. Тогда же было установлено, что перенос ДНК от одних бактериальных клеток к другим осуществляется также в процессе конъюгации бактерий. К настоящему времени трансдукция и конъюгация изучены у многих видов бактерий. В совокупности, на трансформации, трансдукции и конъюгации основывается генетический анализ микроорганизмов. Решающее прямое доказательство генетической роли ДНК было обеспечено разработкой методов генной инженерии, создавшей возможность конструирования рекомбинантных молекул ДНК с заданными свойствами. К настоящему времени возможности генной инженерии показаны на примере клонирования многих генов самых различных организмов. Что касается косвенных доказательств, то они известны очень давно и их несколько. Для ДНК характерна специфичность локализации в клетках, поскольку она обнаруживается только в ядрах клеток (хромосомах), митохондриях (у животных) и хлоропластах (у растений). У многих микроорганизмов ДНК локализована только в ядерной области (нуклеоиде) или в цитоплазме в виде плазмид. Для организмов каждого вида характерно определенное количество ДНК на клетку (табл. 10). Таблица 10 Количество нуклеотидов в геномах различных организмов*
*1000 пар оснований = 617 500 дальтон
Данные, которые приведены в табл. 10, показывают, что, начиная с вирусов, содержание ДНК прогрессивно повышается у бактерий, а затем и у позвоночных и, таким образом, находится в прямой связи со сложностью организации и поведения организмов. Известны, однако, исключения. Для организмов каждого вида характерно то, что количество ДНК в соматических (диплоидных) клетках является вдвое большим, чем в половых (гаплоидных). Будучи постоянным на клетку у организмов всех видов, содержание ДНК не подвержено влиянию со стороны физиологических факторов, включая пол и старение, а также неблагоприятных воздействий, например, голодания, повышения или понижения температуры. Косвенными доказательствами генетической специфичности ДНК являются также данные о способности искусственно синтезированных аналогов азотистых оснований ДНК вызывать наследственные изменения клеток вследствие прямого включения их в ДНК клеток и данные о том, что мутагенный эффект УФ-излучения тесно связан со спектром поглощения его молекулами ДНК.
Химия и структура ДНК
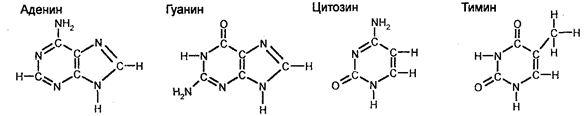
Молекулы ДНК являются линейными макромолекулами, представляющими собой длинные двойные цепи (тяжи) полимеров, составленных из мономеров, получивших название нуклеотидов (малых органических молекул) и являющихся строительными блоками ДНК. Электронномикроскопическое изображение ДНК показано на рис. 106. У всех живых существ макромолекулы ДНК построены по одному и тому же плану. Они слагаются в основном из одних и тех же нуклеотидов, каждый из которых содержит по одной молекуле фосфорной кислоты и сахара, а также одно из четырех азотистых оснований — аденин, гуанин, цитозин или тимин. Аденин и гуанин являются пуриновыми основаниями, тогда как тимин и цито- зин — пиримидиновыми. Пурины и пиримидины называют основаниями по той причине, что в кислой среде они способны присоединять к себе ион H'1'. Пиримидины являются производными шестичленного пиримидинового кольца, тогда как пурины представляют основания, у которых второе пятичленное кольцо слито с шестичленным кольцом.
Сахаром в ДНК является 2-дезокси-D-рибоза, отличающаяся от глюкозы тем, что в ее молекуле не 6, а 5 атомов углерода, т. е. является пятиуглеродным сахаром (пентозой). Особенностью этого сахара является также то, что он имеет атом водорода (Н), присоединенный к одному (специфическому) из атомов углерода, но не гидроксильную группу. Следовательно, этот сахар представляет собой дезоксирибозу, т. к. он является рибозой, лишенной кислорода. Сахарофосфат соединяется с азотистым основанием посредством р-гликозидной связи. Основание прикрепляется к I положению де-зоксирибозы. Структура, образованная соединением азотистого основания и сахара, носит название нуклеозида. Таким образом, химическими группами, которые образуют ДНК, являются пуриновые и пиримидиновые азотистые основания (аденин, гуанин, тимин и цитозин), сахар (2-дезокси-D-рибоза) и фосфорная кислота. РНК характеризуется такой же структурой, как и ДНК. Однако в отличие от ДНК в РНК сахаром является рибоза с кислородом, представляющая собой сахар с 5 атомами углерода, к одному из которых прикреплена 2'-гидроксильная группа (-ОН). Кроме того, в РНК тимин не имеет метильной группы и является урацилом, т. е. в РНК тимин заменен на урацил, также являющийся пиримидиновым основанием. Нуклеиновые кислоты называют кислотами по той причине, что их фосфатные группы освобождают в растворах ионы водорода. Для состава ДНК характерны закономерности, известные в качестве правил А. Чаргаффа, а именно: 1. Сумма нуклеотидов, содержащих пуриновые азотистые основания, равна сумме нуклеотидов, содержащих пиримидиновые азотистые основания
2. Содержание аденина равно содержанию тимина, а гуанина — цитозину ( 3. 4. ДНК из разных источников может иметь различия, обусловленные в одних случаях преобладанием аденина над гуанином и тимина над питозином (А + Т > Г + Ц), в других случаях — преобладанием гуанина и цитозина над аденином и тимином (Г + Ц > А + Т).
Данные о нуклеотидном составе оснований ДНК разных организмов приводятся в табл. 11. Таблица 11
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-12-12; просмотров: 455; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.108 (0.01 с.) |

 Однако изучение других генетических систем, в частности микроорганизмов, показало, что половая репродукция не является единственным путем, при котором осуществляется объединение, расщепление и рекомбинация генетических структур, происходящих от исходных (родительских) организмов. Эти процессы могут проходить и при других формах генетического обмена. У микроорганизмов (Е. coli) бактериальных вирусов (фагов) и микроскопических грибов такими формами генетического обмена являются трансформация, конъюгация и трансдукция. Общим для них в сравнении с половой репродукцией высших организмов является то, что они приводят к объединению в одной клетке родительских генов и обеспечивают их расщепление и рекомбинацию, т. е. являясь альтернативами половой репродукции, представляют собой системы рекомбинации. Поэтому генетический анализ основывается и на таких системах рекомбинации. Больше того, использование этих систем рекомбинации привело к повышению разрешающей способности генетического анализа в гигантских размерах, ибо появилась возможность оперировать с огромным количеством организмов в потомстве, а также легко осуществлять тесты комплементации, а это позволило не только создать генетические карты ряда организмов (Е. coli, В. subtilis, фаги, низшие грибы), но и изучить тонкое строение их генов.
Однако изучение других генетических систем, в частности микроорганизмов, показало, что половая репродукция не является единственным путем, при котором осуществляется объединение, расщепление и рекомбинация генетических структур, происходящих от исходных (родительских) организмов. Эти процессы могут проходить и при других формах генетического обмена. У микроорганизмов (Е. coli) бактериальных вирусов (фагов) и микроскопических грибов такими формами генетического обмена являются трансформация, конъюгация и трансдукция. Общим для них в сравнении с половой репродукцией высших организмов является то, что они приводят к объединению в одной клетке родительских генов и обеспечивают их расщепление и рекомбинацию, т. е. являясь альтернативами половой репродукции, представляют собой системы рекомбинации. Поэтому генетический анализ основывается и на таких системах рекомбинации. Больше того, использование этих систем рекомбинации привело к повышению разрешающей способности генетического анализа в гигантских размерах, ибо появилась возможность оперировать с огромным количеством организмов в потомстве, а также легко осуществлять тесты комплементации, а это позволило не только создать генетические карты ряда организмов (Е. coli, В. subtilis, фаги, низшие грибы), но и изучить тонкое строение их генов.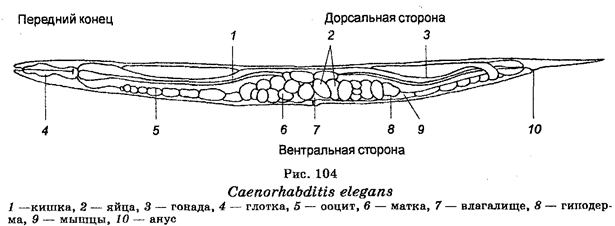
 Классический генетический анализ используют в генетике растений и животных, а также их культивируемых клеток. Однако по отношению к высшим организмам тех видов, которым присуще длительное время между генерациями и малое количество потомства на пару, он либо невозможен, либо очень затруднен. Из-за невозможности классического генетического анализа организмов ряда видов изучение их наследственности проводят с помощью других методов. Например, для изучения наследственности человека используют метод родословных (генеалогический анализ), цитогенетический, популяционный, близнецовый и другие современные методы (см. гл. XIII).
Классический генетический анализ используют в генетике растений и животных, а также их культивируемых клеток. Однако по отношению к высшим организмам тех видов, которым присуще длительное время между генерациями и малое количество потомства на пару, он либо невозможен, либо очень затруднен. Из-за невозможности классического генетического анализа организмов ряда видов изучение их наследственности проводят с помощью других методов. Например, для изучения наследственности человека используют метод родословных (генеалогический анализ), цитогенетический, популяционный, близнецовый и другие современные методы (см. гл. XIII).
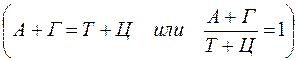
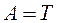 или
или  и
и  или
или 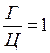 ).
).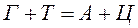 или
или