Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Глава 19 сравнительная психофизиологияСодержание книги
Поиск на нашем сайте Сравнительная психофизиология – наука, нацеленная на установление закономерностей и выявление различий в структурной и функциональной организации мозга, поведении и психике у животных разных видов, в том числе и у человека. Сравнительный метод позволяет оценить филогенетическую близость видов и, тем самым, способствует разработке представлений об эволюции мозга, психики и поведения. Различия между животными разных видов, а также между животными и людьми многообразны. И если морфологические и поведенческие различия между ними, как правило, очевидны, то в том, что касается психики, субъективного мира разных животных подобные заключения оказываются довольно общими и слабо аргументированными, поскольку трудно определить, в чем же заключаются эти различия, а тем более установить их причину. Как известно, существовало два разных подхода к изучению психического. В первом случае для описания субъективного мира животного наблюдатель использовал аналогию с собственным «внутренним миром», что, помимо субъективизма (отсутствия общепризнанной определенности), придавало психике животных антропоморфные черты. Примером такого рода могут служить описания, в которых даже действия беспозвоночных связываются с чувствами, рассудком и свободой воли (Romanes, 1883 [цит. по: Кэндел, 1980, с. 15]), а также утверждения, что «чувства и интуиция, различные эмоции и способности, такие, как любовь, память, внимание, любопытство, подражание, благоразумие и т.д., составляющие предмет гордости человека, в зачаточном, а иногда и хорошо развитом виде могут быть обнаружены у более низкоорганизованных животных» (Darwin, 1871 [цит. по: Кэндел, 1980, с. 14]). Напротив, «объективная» психология по сути отвергала категорию психического, утверждая, что можно «написать труд по психологии..., ни разу не использовав такие термины, как «сознание», «психические состояния», «переживания», проверяемые интроспективно, «образы» и т.п. Все можно описать в понятиях стимулов и реакций, формирования навыков, интеграции этих навыков и т.д.» [Watson, 1913, с. 167]. Оба этих подхода, хотя и в модифицированном виде (особенно второй), в силу развития методов исследования мозга и поведения, – сохранились и до настоящего времени в виде бытовой психологии и «классической» нейрофизиологии, в которой подразумевается, но не всегда эксплицируется, что изучение строения и активности мозга позволит до конца объяснить поведение. Проблемы, связанные с определением психического и поиском адекватных методов его изучения, до сих пор однозначно не решены, что определяет существование разнообразных подходов к их решению (см. гл. 14). ПОЯВЛЕНИЕ ПСИХИЧЕСКОГО Наиболее принятой в настоящее время точкой зрения является та, что психическое – атрибут живых систем, а неживая природа, включая созданные людьми сложные устройства, психикой не обладает. Это важное отличие живых систем от неживых отмечено П. Милнером в «Физиологической психологии»: «Изобретатель даже самой сложной самоорганизующейся машины может, на свое счастье, не беспокоиться о том, чтобы обеспечить своему изобретению непосредственное осознание его входного сигнала, которое, по-видимому, свойственно животным (по крайней мере, мне самому). Он может сделать машину, которая будет сортировать предметы в соответствии с длиной волны отражаемого ими света, но он не может указать на какое-то одно из реле или транзисторов и сказать, что когда через него проходит ток, машина, скажем, «ощущает» желтый цвет. Можно сделать машину, которая будет взвизгивать и подпрыгивать в присутствии определенных раздражителей, и даже такую, которая будет запоминать полученный опыт, чтобы избегать подобных воздействий в будущем, но ее изобретатель не станет утверждать, что вложил в нее способность чувствовать то, что мы называем болью» [Милнер, 1973, с. 14]. Но, признавая появление психики у животных, некоторые авторы, однако, отделяют ее от них, рассматривают психическое как некую отдельную реальность, «ментальный мир» [Popper, Eccles, 1977]. Этот мир может взаимодействовать с мозгом высокоорганизованных животных. Таким образом, если для П. Милнера «осознание» и «ощущение» возникают в самой живой системе, в мозге животного, то, с точки зрения Дж. Экклса [Eccles, 1992], мозг – лишь «место встречи» или «устройство», с помощью которого организм имеет возможность взаимодействовать с «ментальным миром». Несмотря на существующее разнообразие мнений о моменте появления психики (все живое обладает психикой; психика только у животных; только у животных, обладающих нервной системой; только у млекопитающих), наблюдается сходство мнений по поводу того, что появление в эволюции человека явилось началом кардинальных изменений в соотношениях организма со средой и сопровождалось изменениями в психике. Как бы ни определялось психическое (психические процессы и состояния, субъективный мир, индивидуальный опыт), пропасть между инфузорией и человеком – безгранична (рис. 19.1): в своем окружении (рис. 19.1 А, слева) инфузория, по видимому, «выделяет» лишь те немногие объекты, которые для нее витально значимы, например пища (рис. 19.1 А, справа), и, напротив, используя накопленный многообразный опыт поколений и общественное производство, люди оказались способны создавать дворцы, намного превышающие потребности одного человека или семьи в укрытии, и произведения искусства – для удовлетворения «эстетических чувств» (рис. 19.1 Б).
Рис. 19.1. А. Среда обитания инфузории (слева). Весь субъективный мир инфузории (справа) [Uexkull, 1957]. Б. Частичка мира человека Вне зависимости от уровня эволюционного развития, по каким бы критериям сложности ни проводилось сравнение ныне живущих животных, придется согласиться, что «все животные, от наиболее просто организованных до самых сложных, вписаны в их уникальные миры одинаково полно. Простой мир соответствует простым животным, а хорошо структурированный мир (well-articulated world) – сложным» [Uexkull, 1957, р. 11]. Усложнение субъективного мира и принципиальное увеличение приспособительных потенций в эволюционном ряду оказались обусловлены появлением и развитием нервной системы. Значительно большие, даже по сравнению с другими приматами, возможности приспособления человека к действию многообразных и сильных по интенсивности неблагоприятных факторов среды тоже, главным образом, связаны с особенностями организации его мозга. Безусловно, важным оказалось и другое, а именно, что структурные изменения мозга, обусловившие появление специфически человеческих особенностей последнего, произошли у предшественников человека, к этому времени уже «приобретших» плаценту и молочные железы, видоизменененные кости таза, стоп и кистей рук, особенное строение артикуляционного аппарата. Все это в комплексе и позволило человеку занять вершину эволюционной пирамиды. Рассмотрим эволюцию видов и структурной организации мозга для того, чтобы полнее представить картину эволюции психического и выяснить, когда и каким образом могли происходить эти изменения. ЭВОЛЮЦИЯ ВИДОВ Современные данные об эволюции животных указывают на расхождение линий эволюции и развитие параллельных линий (рис. 19.2 А), в том числе и среди млекопитающих (рис. 19.2 Б), и среди приматов (рис. 19.2 В). Это, в частности, означает, что сравнение строения мозга у грызунов, хищных и приматов или же у гориллы, шимпанзе и человека является исследованием не филогенетической, а адаптивной эволюции [Батуев, 1979], поскольку развитие шло в параллельных, независимых друг от друга линиях от общего, в каждом из ранее названных примеров своего, предшественника. Выводы о филогенетической эволюции видов и мозговых структур возможны лишь на основе анализа в последовательных рядах. Например, линия, ведущая к человеку, схематично может быть представлена следующим образом: рептилиеподобные млекопитающие – однопроходные – сумчатые – насекомоядные – обезьяны – приматы – большие обезьяны – африканские человекообразные обезьяны (рис. 19.2 Б, В), хотя и очевидно, что ныне живущие виды могут в той или иной степени отличаться от своих исходных форм – вымерших предков. Прежде чем перейти к краткому описанию основных структурных различий мозга филогенетически разных животных и выяснению эволюционных закономерностей в развитии мозга, следует остановиться на вопросе о том, что является источником многообразия признаков, с которым имеет дело естественный отбор, и в результате каких «событий» появляются новые таксоны. Считается, что процессы развития находятся под генетическим контролем и что эволюцию следует рассматривать как результат изменений в генах, регулирующих онтогенез. Одним из оснований для заключения о том, что морфогенез, а тем самым и морфологическая эволюция (появление новых групп организмов) связаны не столько с изменениями структурных генов, сколько с изменениями регуляторных генов, управляющих генетической программой онтогенеза, являются данные, свидетельствующие о том, что морфологическая эволюция слабо коррелирует с молекулярной эволюцией (эволюцией белков). Так, например, лягушки за 150 млн. лет своей истории почти не изменились ни анатомически, ни по образу жизни, в то время как диапазон морфологического разнообразия у млекопитающих велик: от летучих мышей до китов, от слонов до человека. Несмотря на «морфологический консерватизм» лягушек, аминокислотные последовательности их белков подверглись значительным эволюционным изменениям. И, наоборот, человек и шимпанзе быстро дивергировали и многие систематики относят их к разным семействам, однако аминокислотные последовательности их белков на 99% одинаковы [Рэфф, Кофмен, 1986]. Другими аргументами в пользу морфогенеза за счет регуляторных генов служат данные об отсутствии корреляции между сложностью организации животного и величиной генома, наблюдаемые различия в величине генома у близкородственных животных, которые имеют сходную морфологическую сложность, а также то, что количество генов в геноме гораздо выше количества экспрессируемых (функционирующих) генов.
Рис. 19.2. А. Филогенетическое дерево Metazoa [по Рэфф, Кофман, 1986]. Пунктирная линия – условная граница между таксонами с элементарной сенсорной психикой и таксонами с перцептивной психикой [по Фабри, 1993]
Б. Эволюционное дерево млекопитающих с выделением этапов филогенетической эволюции и уровней адаптивной эволюции [Батуев, 1979]. В. Филогенетическое дерево высших обезьян [Povinelli, Preuss, 1995] Регуляторные гены являются «переключателями» между альтернативными путями развития, и количество их относительно невелико. Они функционируют на протяжении всего процесса развития, управляя процессом онтогенеза тремя различными способами: 1) регулируя время наступления тех или иных событий; 2) делая выбор из двух возможностей, тем самым определяя судьбу клеток и частей зародышей и 3) интегрируя экспрессию структурных генов и обеспечивая создание стабильных дифференцированных тканей. Поскольку регуляторный ген способен влиять на скорость роста какой-то определенной структуры, он контролирует как сроки появления, так и размеры данной структуры. А поскольку онтогенез слагается из связанных между собой процессов, т.е. формирование каждой отдельной структуры зависит как во времени, так и в пространстве от формирования других структур, изменения в сроках возникновения одного морфогенетического события могут иметь глубокие последствия, изменяя многие дальнейшие зависящие от него ступени онтогенеза [Рэфф, Кофмен, 1986]. Важно отметить, что модификации генетических регуляций приводят к возникновению новых структур, в том числе и разных «конструкций» мозга, а тем самым, и типов поведения, открывающих новые адаптивные возможности. Очевидно, что бифуркации на всех схемах, представленных на рис. 19.2, связаны именно с такими процессами появления новых признаков. В дальнейшем, после бифуркаций, изменения носят характер адаптивной эволюции, как это, например, показано на рис. 19.2 Б в ряду млекопитающих. Такой тип морфогенеза может объяснить «скачки» в развитии живого, связанные как с возникновением нервной системы и появлением психики, так и с формированием сложноорганизованного мозга человека с его отличной от других животных психикой, а также то, почему, даже приложив неимоверные усилия и создав все условия, не удастся «превратить» шимпанзе в человека, енота «заставить» говорить, а кролика – охотиться на слонов. Наряду с зоологической, существует и психологическая классификация животных. Одной из них является классификация К.Э. Фабри, для создания которой он использовал представления А. Н. Леонтьева о «психике как форме отражения, позволяющей животному организму адекватно ориентировать свою активность по отношению к компонентам среды» [Фабри, 1993, с. 7], а в качестве уровней развития психики выделил уровень элементарной сенсорной психики и уровень перцептивной психики, граница между которыми проходит на уровне членистоногие, головоногие и хордовые (см. пунктирную линию на рис. 19.2 А), хотя оказалось, что не все крупные таксоны укладываются в эти рамки. Для высших позвоночных в этой классификации имеется стадия интеллекта, при том, что отмечаются принципиальные различия между интеллектом животных и людей, заключающиеся в том, что мышление животных всегда имеет конкретно-чувственный характер и осуществляется через действия с предметами. Для наиболее часто используемых определений психики (а именно, психика как психические процессы и состояния – субъективный мир – индивидуальный опыт) детальных психологических классификаций животного мира пока не создано. ЭВОЛЮЦИОННЫЕ ПРЕОБРАЗОВАНИЯ МОЗГА Строение мозга у животных разных видов разнообразно. И хотя, как следует из рис. 19.3, родственные виды, например среди ракообразных или млекопитающих, имеют общие черты в строении мозга, между собой они мало похожи. Именно с появлением нервной системы организмы получили возможность быстрой адаптации к быстро меняющимся факторам среды, что, безусловно, давало преимущества в борьбе за существование. У предков млекопитающих, как и у современных рептилий, кора больших полушарий была очень слабо дифференцирована [Батуев, 1979]. Но на пути от рептилиеподобных предков млекопитающих до ныне живущих млекопитающих произошло значительное увеличение коры мозга по сравнению с другими структурами мозга (рис. 19.4), изменение количества (см. рис. 19.3 В), размеров и формы корковых областей, их модульного строения и связей [Kaas, 1987], произошло увеличение переднего мозга и, как предполагается [McLean, 1993; Povinelli, Preuss, 1995], перемещение главенствующей роли от стриарного комплекса сначала к лимбической системе, а затем к неокортексу и особенно к префронтальной области коры (см. рис. 19.3 Б). Важной чертой эволюции млекопитающих является уже упомянутое увеличение неокортекса, при этом степень этого увеличения отличает приматов от остальных млекопитающих, а человека – от остальных приматов. В частности, соотношение площади неокортекса у мыши, макаки и человека составляет 1: 100: 1000 соответственно [цит. по: Rakic, 1995], а соотношение объемов коры мозга и спинного мозга у крыс и у человека – 31: 35 и 77: 2, соответственно (см. рис. 19.4) [Swanson, 1995]. Такая экспансия неокортекса, как полагают, связана с модификацией регуляторных генов, определяющих митотическое деление клеток вентрикулярного (пролиферативного) слоя коры [Rakic, 1995]. При этом, по-видимому, кора развивалась под влиянием внутрикорковых или непрямых таламических подкорковых влияний [Krubitzer, 1995]. Такое развитие коры привело к возникновению новых мощных приспособительных функций [Innocenti, Kaas, 1995]. Можно предположить, что именно это явилось основой такого свойства нервной системы, которое У. Матурана обозначил как способность нервной системы «взаимодействовать со своими собственными внутренними состояниями, как если бы те были независимыми сущностями» [Матурана, 1995, с. 104]. В таком «взаимодействии» нервной системы со своими собственными состояниями может лежать ключ и к пониманию онтологической сути субъективных состояний – «переживаний», «мыслей», «образов», – а также языка и мышления.
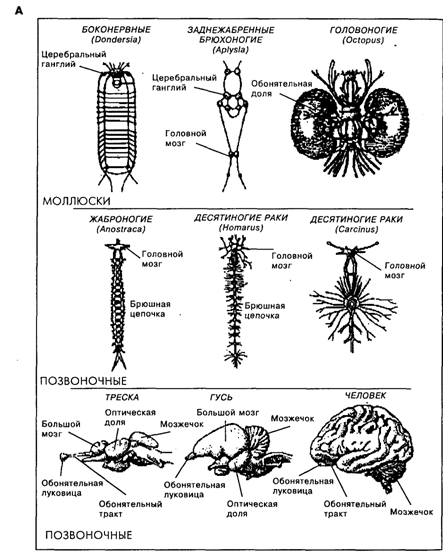
Рис. 19.3. А. Схематичное представление мозга животных, относящихся к линиям моллюсков, членистоногих и хордовых (масштабы разные)[по Кэндел, 1980] Б. Условное представление триединой эволюции трех основных мозговых ансамблей, приведших к образованияю переднего мозга у людей и у других развитых млекопитающих [McLean, 1993]. В. Корковые поля у примитивных млекопитающих (еж), у млекопитающих с незначительно более развитой нервной системой (белка) и у двух млекопитающих с умеренно развитым мозгом (кошка и совиная обезьяна) [Kaas, 1987]
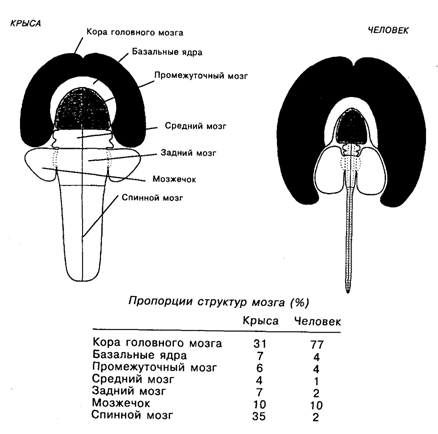
Рис. 19.4. Сравнение размеров разных отделов цетральной нервной системы крысы и человека. Пропорции между областями мозга соблюдены [Swonson, 1995] Внутрикорковые процессы рассматриваются рядом авторов в качестве нейронной основы сознания [Маунткасл, 1981; Эделмен, 1981; Eccles, 1992]. Так, например, в концепциях В. Маунткасла и Дж. Эделмена сознание возникает на основе повторного входа «сигнализации» (см. гл. 11), при этом, согласно одним взглядам, данный процесс происходит в модульных распределенных сетях [Маунткасл, 1981], а согласно другим – в вырожденной группе нейронов [Эделмен, 1981]. По мнению же Дж. Экклса, сознание – побочный эффект, возникший в эволюции неокортекса, наблюдающийся только у млекопитающих и сводящийся к взаимодействию мозга и ментального мира в множестве пресинаптических окончаний корковых нейронов, которые образуют морфологическую единицу – дендрон [Eccles, 1992].
|
||
|
|
Последнее изменение этой страницы: 2016-09-20; просмотров: 285; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.62 (0.015 с.) |