Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Дифференциальная сенсорная чувствительностьСодержание книги
Поиск на нашем сайте Дифференциальная сенсорная чувствительность основана на способности сенсорной системы к различению сигналов. Важная характеристика каждой сенсорной системы – способность замечать различия в свойствах одновременно или последовательно действующих раздражителей. Различение начинается в рецепторах, но в нем участвуют нейроны всех отделов сенсорной системы. Оно характеризует то минимальное различие между стимулами, которое человек может заметить (дифференциальный или разностный порог). Порог различения интенсивности раздражителя практически всегда выше ранее действовавшего раздражения на определенную долю (закон Вебера). Так, усиление давления на кожу руки ощущается, если увеличить груз на 3% (к гирьке весом в 100 г надо добавить 3 г, а к гирьке весом в 200 г надо добавить 6г). Эта зависимость выражается следующей формулой: dI/1 = const., где I – сила раздражения, dI – ее едва ощущаемый прирост (порог различения), const – постоянная величина (константа). Аналогичные соотношения получены для зрения, слуха и других органов чувств человека. Зависимость силы ощущения от силы раздражения (закон Вебера–Фехнера) выражается следующей формулой: Е = a log I + b, где Е – величина ощущения, I – сила раздражения, а и b – константы, различные для разных модальностей стимулов. Эта формула показывает, что ощущение усиливается пропорционально логарифму интенсивности раздражения. Кроме того, современная психофизиология для оценки силы ощущения использует также методы сенсорного шкалирования, т.е. субъективной оценки человеком силы своего ощущения путем его сравнения с ранее созданным эталоном или набором таких эталонов. Отношение между ощущением и стимулом в этом случае выражается степенной функцией (закон Стивенса), Сравнение логарифмической функции закона Вебера – Фехнера и степенной функции закона Стивенса показало, что в основной, рабочей части диапазона интенсивностей эти функции дают количественно близкие оценки. Ранее говорилось о различении силы раздражителей. Пространственное различение сигналов основано на характере распределения возбуждения в слое рецепторов и в нейронных слоях сенсорной системы. Так, если два раздражителя возбудили два соседних рецептора, то их различение невозможно: они сольются и будут восприняты как единое целое. Необходимо, чтобы между двумя возбужденными рецепторами находился хотя бы один невозбужденный. Временное различение двух раздражений возможно, если вызванные ими нервные процессы не сливаются во времени, а сигнал, вызванный вторым стимулом, не попадает в рефракторный период от предыдущего раздражения. Нейрофизиологической основой временного разрешения являются так называемые циклы возбудимости, или циклы восстановления ответов. О них судят по величине ответа на второй из двух последовательно предъявленных стимулов. При коротких интервалах между стимулами ответа на второй из них может не быть вообще (абсолютный рефракторный период). У человека по поведенческим реакциям этот период может длиться от нескольких десятков до 100 и более миллисекунд. При больших интервалах ответ на второй стимул появляется, но величина его меньше, чем на одиночный стимул (относительная рефрактерность). И, наконец, при еще больших интервалах восстановление второго ответа заканчивается, и он сравнивается с ответом на одиночное раздражение. На временном взаимодействии между последовательными раздражителями основана так называемая «сенсорная маскировка». Она лежит в основе многих сенсорных эффектов и широко используется в психофизиологических экспериментах. Сама маскировка прямо связана с попаданием одного из стимулов в рефракторную фазу цикла возбудимости после первого раздражения. Различают прямую маскировку, при которой тормозится ответ на второй стимул, и обратную маскировку, при которой второй стимул как бы прерывает или мешает обработке информации о первом сигнале. Эффективность как прямой, так и обратной маскировки тем больше, чем короче интервал между стимулом и «маской», а также чем более сходны эти два сигнала по своим свойствам. В качестве «маски» часто используют стимул, состоящий либо из шума, либо из набора хаотично распределенных элементов основного раздражителя. ПЕРЕДАЧА И ПРЕОБРАЗОВАНИЕ СИГНАЛОВ Процессы передачи и преобразования сигналов обеспечивают поступление в высшие сенсорные центры наиболее важной (существенной) информации о сенсорном событии в такой форме, которая удобна для надежного и быстрого анализа. Что следует считать существенной информацией? В разных условиях и ситуациях это понятие может меняться. Однако имеется некоторое общее свойство, которое универсально отличает существенную информацию от несущественной. Это – степень ее новизны. Ясно, что новые события при прочих равных условиях информационно важнее для организма, чем привычные. Поэтому эволюционно было выработано свойство прежде всего и быстрее всего передавать в мозг и перерабатывать информацию об изменениях в сенсорной среде. Эти изменения могут быть как временными, так и пространственными. Среди пространственных преобразований выделяют изменение представительства размера или соотношения разных частей сигнала. Так, в соматосенсорной и зрительной системах на корковом уровне значительно искажаются геометрические пропорции представительства отдельных частей тела или частей поля зрения. В зрительной коре резко расширено представительство информационно наиболее важной центральной ямки сетчатки, ответственной за детальное «поточечное» описание изображения при относительном сжатии проекции периферии поля зрения («циклопический глаз»). В соматосенсорной коре также преимущественно представлены наиболее важные для тонкого различения и организации поведения зоны тела – кожа пальцев рук и лица («сенсорный гомункулюс»; см. гл. 4). Различные проекционные корковые зоны, например зрительной системы (а их насчитывают несколько десятков), отличаются характером ретинотопии, т.е. представительства разных частей сетчатки. Так, имеются зоны, в которых представлен только центр сетчатки, или, наоборот, только ее периферия. Это связано со специфическим участием каждой из зон в зрительном восприятии: обслуживанием преимущественно предметного зрения или обработкой информации о движениях стимулов в поле зрения. Для временных преобразований информации во всех сенсорных системах типично сжатие, или временная компрессия сигналов: переход от длительной (тонической) импульсации нейронов на нижних уровнях системы к коротким (фазическим) пачечным разрядам нейронов высоких уровней. Ограничение избыточности информации. Зрительная информация, идущая от фоторецепторов, могла бы очень быстро насытить все информационные резервы мозга. Примерно то же самое, пусть несколько медленнее, могло бы происходить при работе и других сенсорных систем. Огромная избыточность первичных сенсорных сообщений, идущих от рецепторов, ограничивается путем подавления информации о менее существенных сигналах. Менее важно во внешней среде то, что неизменно либо изменяется медленно во времени и в пространстве (см. ранее). Например, на сетчатку глаза или на кожу длительно действует неизменный стимул большого размера. Для того чтобы постоянно не передавать в мозг информацию от всех возбужденных рецепторов, сенсорная система пропускает в мозг сигналы только о начале, а затем об окончании раздражения, причем до коры доходят сообщения только от рецепторов, которые лежат по контуру возбужденной области. КОДИРОВАНИЕ ИНФОРМАЦИИ Кодированием называют совершаемое по определенным правилам преобразование информации в условную форму – код. В сенсорной системе сигналы кодируются двоичным кодом, т.е. наличием или отсутствием электрического импульса в тот или иной момент времени [Сомьен, 1975]. Такой способ кодирования крайне прост и устойчив к помехам. Информация о раздражении и его параметрах передается в виде отдельных импульсов, а также групп, или «пачек» импульсов. Амплитуда, длительность и форма каждого импульса одинаковы, но количество импульсов в пачке, частота их следования, длительность пачек и интервалов между ними, а также временной «рисунок» (pattern) пачки различны и зависят от характеристик стимула. Сенсорная информация кодируется также числом одновременно возбужденных нейронов и их расположением в нейронном слое. Особенности кодирования в сенсорных системах. В отличие от телефонных или телевизионных кодов, которые декодируются восстановлением первоначального сообщения в исходном виде, в сенсорной системе подобного декодирования не происходит. Еще одна важная особенность нервного кодирования – множественность и перекрытие кодов. Так, для одного и того же свойства сигнала (например, его интенсивности) сенсорная система использует несколько кодов: частота и число импульсов в пачке, число возбужденных нейронов и их локализация в слое. В коре мозга сигналы кодируются также последовательностью включения параллельно работающих нейронных каналов, синхронностью ритмических импульсных разрядов возбужденных нейронов, изменением их числа. В коре одним из основных используемых способов становится позиционное кодирование. Оно заключается в том, что какой-то признак раздражителя вызывает возбуждение определенного нейрона или небольшой группы нейронов, расположенных в определенном месте нейронного слоя. Например, возбуждение небольшой локальной группы нейронов зрительной коры означает, что в определенной части поля зрения появилась световая полоска определенного размера и ориентации. Возбуждение определенных нейронов височной коры сигнализирует о появлении в поле зрения знакомого лица. Для периферических отделов сенсорной системы типично временное кодирование признаков раздражителя, а на высших уровнях происходит переход к преимущественно пространственному (в основном позиционному) коду. ДЕТЕКТИРОВАНИЕ СИГНАЛОВ Детектированием называют избирательное выделение сенсорным нейроном того или иного признака раздражителя, имеющего поведенческое значение. Осуществляют такой анализ нейроны-детекторы, избирательно реагирующие лишь на определенные свойства стимула. Так, типичный нейрон зрительной коры отвечает разрядом лишь на один из наклонов (ориентацию) световой полоски, расположенной в определенной части поля зрения [Хьюбел, 1990]. При других наклонах той же полоски ответят другие нейроны. Такие нейроны называют детекторами первого порядка, так как они выделяют наиболее простые признаки сигнала. В высших отделах сенсорной системы сконцентрированы детекторы высших порядков, ответственные за выделение сложных признаков и целых образов. Примером могут служить детекторы лиц, найденные в нижневисочной коре обезьян (предсказанные много лет назад Ю. Конорским, они были названы «детекторами моей бабушки»). Многие детекторы формируются в раннем онтогенезе под влиянием внешней среды, а у части из них детекторные свойства заданы генетически. ОПОЗНАНИЕ ОБРАЗОВ Это конечная и наиболее сложная операция сенсорной системы. Она заключается в отнесении образа к тому или иному классу объектов, с которыми ранее встречался организм, т.е. в классификации образов. Синтезируя сигналы от нейронов-детекторов, высший отдел сенсорной системы формирует «образ» раздражителя и сравнивает его со множеством образов, хранящихся в памяти. Опознание завершается принятием решения о том, с каким объектом или ситуацией встретился организм. В результате этого происходит восприятие, т.е. мы осознаем, чье лицо видим перед собой, кого слышим, какой запах чувствуем. Нейрофизиологические механизмы опознания сенсорных образов исследованы пока недостаточно. Опознание часто происходит независимо от изменчивости сигнала. Мы надежно опознаем, например, предметы при различной их освещенности, окраске, размере, ракурсе, ориентации и положении в поле зрения. Это означает, что сенсорная система формирует независимый от изменений ряда признаков сигнала (инвариантный к этим изменениям) сенсорный образ. При опознании сенсорных образов возможны ошибки. Особую группу таких ошибок составляют так называемые «сенсорные иллюзии». Они основаны на некоторых побочных эффектах взаимодействия нейронов, участвующих в обработке сигналов, и приводят к искаженной оценке образа в целом или отдельных его характеристик (размер, соотношение частей и т.п.). Пример иллюзии последнего типа представлен на рис. 3.2.
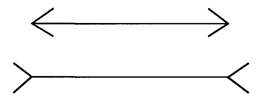
Рис. 3.2. Пример одной из простейших зрительных иллюзий: длина горизонтальных отрезков на верхней и нижней фигурах кажется различной, хотя на самом деле они равны АДАПТАЦИЯ СЕНСОРНОЙ СИСТЕМЫ Сенсорная система обладает способностью приспосабливать свои свойства к условиям среды и потребностям организма. Сенсорная адаптация – это общее свойство сенсорных систем, заключающееся в приспособлении к длительно действующему (фоновому) раздражителю. Существует общая, или глобальная, и локальная, или селективная адаптация. Общая, или глобальная, адаптация проявляется в снижении абсолютной и повышении дифференциальной чувствительности всей сенсорной системы. Субъективно адаптация проявляется в привыкании к действию постоянного раздражителя (например, мы не замечаем непрерывного давления на кожу привычной одежды). Локальная, или селективная, адаптация сводится к снижению чувствительности не всей сенсорной системы, а какой-либо ее части, подвергнутой длительному действию стимула. Так, порог реакции оказывается избирательно повышен для изображения решетки с определенной пространственной частотой (определенным периодом чередования черно-белых полос). Пороги реакций на соседние пространственные частоты при этом не изменяются [Глезер, 1985]. Локальная адаптация часто используется в психофизиологических работах для выявления так называемых «сенсорных каналов», ответственных за обработку сведений о том или ином признаке сигнала. Адаптационные процессы начинаются на уровне рецепторов, охватывая и все нейронные уровни сенсорной системы. Заметная адаптация не развивается только в вестибулои проприорецепторах. По скорости данного процесса все рецепторы делятся на быстро и медленно адаптирующиеся. Первые после развития адаптации практически вообще не сообщают в мозг о длящемся раздражении, у вторых эта информация передается, хотя и в значительно ослабленном виде. Когда действие постоянного раздражителя прекращается, абсолютная чувствительность сенсорной системы восстанавливается. Так, в темноте абсолютная чувствительность зрения резко повышается. В сенсорной адаптации важную роль играет эфферентная регуляция свойств сенсорной системы. Она осуществляется за счет нисходящих влияний со стороны более высоких на более низкие ее отделы. Происходит как бы перенастройка свойств нейронов на оптимальное восприятие внешних сигналов в изменившихся условиях. Кроме того, состояние разных уровней сенсорной системы контролируется также ретикулярной формацией, включающей их в единую систему, интегрированную с другими отделами мозга и организма в целом. Эфферентные влияния в сенсорных системах чаще всего имеют тормозный характер, т.е. приводят к уменьшению их чувствительности и ограничивают поток афферентных сигналов. Общее количество эфферентных нервных волокон, приходящих к элементам какого-либо нервного слоя, как правило, во много раз меньше количества его собственных нейронов. Это определяет важную особенность эфферентного контроля в сенсорных системах: его широкий и диффузный характер. Речь идет об общем снижении чувствительности значительной части нейронного слоя.
|
||
|
|
Последнее изменение этой страницы: 2016-09-20; просмотров: 459; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.62 (0.015 с.) |