Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Посттранскрипционная регуляцияСодержание книги
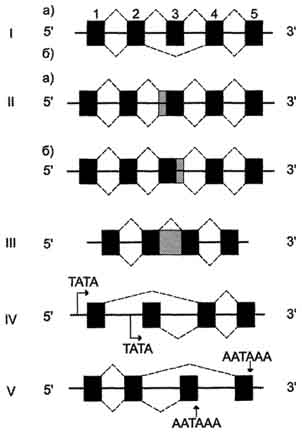
Поиск на нашем сайте В организме животных существенное значение в обеспечении разнообразия белков играет посттранскрипционный процессинг РНК. Основные способы такой регуляции - альтернативный сплайсинг и изменение стабильности РНК. Альтернативный сплайсинг. Установлено, что многие эукариотические гены, будучи транскрибированы, образуют несколько вариантов зрелой мРНК в ходе процессинга (или созревания) первичного транскрипта, имеющего полиэкзонное строение. Возможные варианты сплайсинга РНК представлены на рис. 4-53. Наиболее часто промотор сохраняется на одном из концов транскрипта, а в ходе сплайсинга происходит "вырезание" одного или нескольких экзонов. В других случаях в зрелой мРНК сохраняется часть интрона и включается в состав экзона с 5' или 3'-конца. Сплайсинг может влиять на выбор промотора или участка полиаденилирования. С помощью альтернативного сплайсинга в процессе синтеза антител образуются мембра-носвязанные и секреторные формы антител (рис. 4-54). Так, первоначально В-лимфоциты продуцируют транскрипты, полиаденилированные после второго стоп-кодона, а интрон, в котором имеется первый стоп-кодон, удаляется. В результате синтезируются IgM, связанные с клеточной мембраной, так как мРНК таких клеток содержит на 3'-конце экзон, кодирующий участок полипептидной цепи, состоящий из гидрофобных аминокислот. С помощью этого участка происходит "заякоривание" IgM в мембране. Когда В-лимфоциты превращаются в плазматические клетки, то в
Рис. 4-53. Часто встречающиеся варианты сплайсинга первичных транскриптов РНК. I. Вырезание одного из экзонов: а) синтез белка, содержащего полный набор экзонов (1-5); б) синтез белка, лишённого одного экзона (1, 2,4, 5); II. Сохранение участка интрона: а) с 5'-конца; б) с 3'-конца. III. Сохранение целого интрона. IV. Использование альтернативных промоторов (либо перед экзоном 1, либо перед экзоном 2). V. Использование альтернативных участков полиаденилирования (например, при последовательном сшивании экзонов после экзона 3, а если экзон 3 не прочитывается, то после экзона 4). результате альтернативного сплайсинга образуется мРНК, в которой сохраняется интрон, содержащий первый стоп-кодон. Поэтому происходит более раннее полиаденилирование и исчезает экзон, кодирующий гидрофобный участок молекулы. Синтезируются укороченные молекулы антител, секретируемые в кровь. "Редактирование" РНК. Описан ряд случаев, когда первичная структура мРНК изменяется ("редактируется") после транскрипции. Последовательность нуклеотидов в таких генах одинакова, а транскрибируемая в разных тканях мРНК различается в результате появления в молекуле замен, вставок или выпадений нуклеотидов. Пример "редактирования" РНК - образование апопротеина В (апо-В) в клетках печени и тонкого кишечника (рис. 4-55). Апо-В - основной компонент липопротеинов, участвующих в транспорте триацилглицеринов из этих тканей в кровь. Хотя апопротеин В кодируется одним и тем же геном, вариант белка, образующийся в печени, называют апо-В-100, и он содержит 4563 аминокислотных остатка, тогда как белок, синтезированный в клетках кишечника, состоит из 2152 аминокислот. В гене, кодирующем этот белок, последовательность нуклеотидов в триплете 2153 - САА и шифрует включение в полипептидную цепь остатка глутамина. В клетках кишечника в первичном транскрипте гена азотистое основание - цитозин (С) ко-дона 2153 дезаминируется и превращается в урацил (U). Возникает стоп-кодон - UAA, прекращающий трансляцию мРНК в середине молекулы и приводящий к синтезу укороченного белка. В результате образуется белок (В-48), длина которого составляет 48% от длины белка синтезируемого печенью. Изменение стабильности мРНК. Для того, чтобы участвовать в синтезе белка, мРНК должна выйти из ядра в цитоплазму через ядерные поры. Установлено, что в ядре клеток обычно синтезируется больший набор гетерогенных РНК, чем тот, что выходит в цитоплазму. Многие продукты транскрипции подвергаются расщеплению нуклеазами, а те мРНК, что, транспортируются из ядра в цитоплазму, защищаются от гидролитического разрушения, образуя комплексы с белками. Время жизни эукариотических мРНК значительно больше (t1/2 составляет от нескольких часов до нескольких дней), чем t1/2 мРНК прокариотов, равное нескольким минутам. Очевидно, что стабильность молекул мРНК - фактор, изменение которого влияет на уровень трансляции. Стабилизация мРНК при фиксированной скорости транскрипции приводит к накоплению и увеличению количества образующегося белкового продукта. Продолжительность жизни разных мРНК варьирует в достаточно широких пределах. Некоторые гены кодируют продукт с большой продолжительностью жизни. Так, в ходе транскрипции гена β-глобина образуется мРНК с t1/2, равной примерно 10 ч. Другие гены образуют мРНК с короткой продолжительностью жизни: мРНК, на которых синтезируются факторы роста,
Рис. 4-54. Использование механизмов альтернативного сплайсинга и полиаденилирования в ходе синтеза мембранно-связанных и секреторных lg. Если транскрипт подвергается полиаденилированию после второго стоп-кодона, присутствующего в экзоне гена lgM, то синтезируются белки, у которых на С-конце присутствует гидрофобный домен, обеспечивающий связывание с плазматической мембраной. При стимуляции В-лимфоцитов в клетках осуществляется альтернативный сплайсинг первичного транскрипта, при котором интрон, содержащий первый стоп-кодон, сохраняется. Образуются более короткие мРНК, полиаденилирование которых происходит после первого стоп-кодона.
Рис. 4-55. "Редактирование" мРНК апопротеина В. В ходе транскрипции гена апопротеина В в печени образуется мРНК, служащая матрицей для синтеза белка, состоящего из 4563 аминокислотных остатков. В клетках тонкого кишечника экспрессия того же гена вызывает образование белка, состоящего из 2152 аминокислот. В РНК транскрипте цитозин кодона 2153 - САА превращается в урацил (U), и бозникает стоп-кодон в середине молекулы мРНК. Это приводит к синтезу укороченного белка.
Рисунок 4-56. Зависимость скорости синтеза глобина от концентрации тема. Когда внутриклеточный уровень тема высок, фактор инициации elF2 не фосфорилирован и активен, происходит синтез глобина. Если содержание тема в клетке снижается, фактор инициации фосфорилируется, инактивируется и синтез белка прекращается. имеют t1/2 менее 1 ч. Показано, что поли(А)-фрагмент на 3'-конце мРНК увеличивает продолжительность жизни молекул. Чем длиннее поли(А)-фрагмент, тем больше время жизни мРНК. Описано много примеров регуляции количества синтезирующихся белков за счёт изменения продолжительности функционирования мРНК. Так, стабильность мРНК-матриц для синтеза молекул гистонов сильно зависит от фазы клеточного цикла. В S-фазе гистоны постоянно синтезируются и используются для укладки вновь образованной ДНК в нуклеосомы. Гистоновая мРНК в этот период стабильна в течение нескольких часов. После S-периода, когда ДНК уже не синтезируется, в клетках образуется небольшое количество гистонов, так как они не требуются для формирования нуклеосом. В этот период t1/2 для гистоновой мРНК составляет 10-15 мин. 6. Регуляция трансляции и посттрансляционных
|
||
|
|
Последнее изменение этой страницы: 2016-09-18; просмотров: 588; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.119 (0.008 с.) |