
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Організація генома эукариот.Содержание книги
Поиск на нашем сайте
Організмений рівень генетичної інформації представлений геномом і генотипом. Геном видоспецифичен і представляє той необхідний набір генів, який забезпечує формування видових характеристик організмів в онтогенезі. При статевому розмноженні в процесі запліднення об'єднуються геноми двох батьківських гамет, утворюючи генотип нового організму. Всі соматичні клітки такого організму мають подвійний набір генів, отриманих від обох батьків. Генотип людини сформувався еволюційно. Загальна еволюція генотипу эукариот пов'язана з прогресивним збільшенням кількості ДНК. Серед можливих механізмів збільшення генома виділяють полиплоидизацию і ампліфікацію. Поліплоїдія (збільшення кількості ДНК і хромосом, кратне гаплоидному набору) збільшує дозу всіх генів і утворює надлишок генетичного матеріалу, який потім змінюється в результаті ті мутацій і відбору. В процесі еволюції полиплоидизация супроводжувалася переходом в гаплоидное стан. Другий механізм – ампліфікація (утворення копій ділянок ДНК). Певне значення в збільшенні і перетворенні генома мали хромосомні перебудови (делеции, дупликации, транслокации). Сучасні уявлення про геном людини. Організація генома кожного эукариотического виду є послідовною ієрархією елементів: нуклеотидов, кодонів, доменів, генів з міжгенними ділянками, складних генів, плечей хромосом, хромосом, гаплоидного набору разом з внехромосомной і внеядерной ДНК. У еволюційному перетворенні генома кожен з цих ієрархічних рівнів міг поводитися абсолютно дискретно (змінюючись, комбінуючись з іншими і так далі). Наші уявлення про геном людини — обширна область генетики людини, що включає, щонайменше, поняття «інвентаризації» генів, груп зчеплення, картирования генів (локалізація), секвенирования всієї ДНК (генів, їх мутацій і хромосом в цілому), мейотических перетворень, функціонування окремих генів і їх взаємодій, інтеграції структури і функції генома в цілому. На вирішенні всіх цих питань була зосереджена обширна багаторічна міжнародна програма «Геном людини» (з 1990 по 2003 р.). Головним напрямом робіт були послідовні секвенирование ділянок генома і їх «стикування». Успішні розробки в цій області додали програмі клініко-генетичний аспект (табл.9).
Табл.9. Клінічні додатки відомостей про геном людини
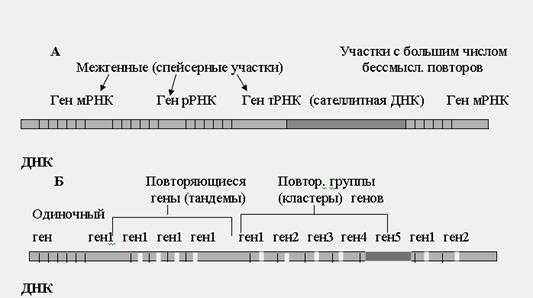
Систематичне вивчення генома людини фактично почалося із застосування аналізу Менделя спадкових ознак людини (початок XX століття). Генеалогічний метод увійшов тоді до широкої практики, і крок за кроком почав накопичуватися матеріал по «інвентаризації» дискретних спадкових ознак людини, але цей процес поступово сповільнювався (за 50 років було відкрито не більше 400 менделирующих ознак і 4 групи зчеплення), можливості клинико-генеалогического методу в чистому вигляді були вичерпані. Бурхливий прогрес цитогенетики людини, біохімічної генетики і особливо генетики соматичних кліток в 60-х роках в комплексі з генеалогічним підходом поставив вивчення генома людини на нові теоретичні основи і високий методичний рівень. Виявлення нових менделирующих ознак людини почало швидко просуватися, особливо на біохімічному і імунологічному рівні, з'явилися можливості вивчення зчеплення і локалізації генів. Особливий імпульс вивченню генома людини додали молекулярно-генетичні методи, або технологія генної інженерії (70-і роки). Процес пізнання генома поглибився до виділення гена в чистому вигляді і його секвенирования. На відміну від класичної, в новій генетиці змінився підхід до аналізу генів. У класичній генетиці послідовність була наступною: ідентифікація менделируюшего ознаки - локалізація гена в хромосомі (або групі зчеплення) - первинний продукт гена - ген. У сучасній генетиці став можливим і зворотний підхід: виділення гена - секвенирование - первинний продукт, у зв'язку з чим був введений новий термін для визначення такого напряму досліджень: «зворотна генетика» або «генетика навпаки». Продовжуються вдосконалення молекулярно-генетичних методів і, що не менш важливий, їх автоматизація. У США і Великобританії були розроблені і упроваджені автоматичні прилади по секвенированию геномів. Їх назвали геномотронами. У них здійснюється до 100 000 полимеразных реакцій в годині Це означає, що протягом тижня може бути просеквенирован ділянка (або ділянки) завдовжки в декілька мільйонів пар нуклеотидов. Велику роль в розшифровці генома людини грають обчислювальна техніка і інформаційні системи. Завдяки ним вирішуються питання накопичення інформації (бази даних) з різних джерел, зберігання її і оперативного використання дослідниками з різних країн. Характеристика генома людини. Структура генома. Геномом (від слів ген + хромосома) називається сукупність всієї спадкової інформації організму (всіх генів і міжгенних послідовностей нуклеотидов). Розмір генома людини складає 3 мільярди пар підстав. Кожна з 23 пар хромосом містить окрему лінійну двунитевую молекулу ДНК. Розмір ДНК в найбільшій хромосомі 1 (хромосоми нумерують за розміром) - 250 мільйонів пар нуклеотидов, а в найменшій - 47 мільйонів. У кожній клітці людини близько 22-25 тисяч пар генів, але точне їх число поки не відоме. У генах записана інформація про структуру молекул РНК: матричною (кодуючої білки), рибосомной, транспортною і деяких інших видів так званої некодуючою РНК. Середній розмір гена в хромосомі складає близько 50 тисяч пар нуклеотидов. Найкоротші гени містять всього два десятки букв-нуклеотидов, наприклад, гени эндорфинов - білків, що викликають відчуття задоволення. Гени інтерферонів - білків, що захищають людину від вірусних інфекцій, мають розмір близько 700 нуклеотидов. Щонайдовший ген, що кодує один з білків м'язів, - миодистрофин, містить 2,5 мільйона букв. У примітивних організмів, таких як бактерії, гени займають близько 80-90% всієї ДНК. У людини на гени доводиться, мабуть, не більше 5% нуклеотидных послідовностей. Решту ДНК раніше називали надмірною, але з часом стало ясно, що вона виконує важливі функції, зокрема містить інформацію про те, як, в якому порядку повинні включатися гени. Біля третини генома доводиться на послідовності різної довжини, що повторюються. На початку і в кінці гена знаходяться регуляторні послідовності, які визначають, в яких тканинах, на яких етапах розвитку і при яких зовнішніх або внутрішніх (наприклад, гормональних) сигналах працюватиме даний ген. Регуляторні послідовності знаходяться не тільки поряд з генами, але і в ділянках, що містять так звану реторовирусную ДНК - залишки ретровірусних геномів, які колись вбудувалися в геном людини і переходять в його складі з покоління в покоління. Ретровіруси належать до широкої групи генетичних елементів, реплицирующихся за допомогою зворотної транскрипції. Деякі ретровіруси не пов'язані з якою-небудь хворобою, тоді як інші дуже патогенні, такі як вірус гепатиту В і вірус імунодефіциту людини. Ретровіруси інфікують різноманітні види хребетних, від риб до людини. Під час реплікації ретровірус копіює свій РНК-геном в ДНК, використовуючи кодований вірусним геномом фермент - зворотну транкриптазу (ревертазу). Вірусна ДНК вбудовується в господарські хромосоми за допомогою іншого вірусного ферменту - интегразы. Якщо вірусний геном вбудовується в господарські гени, це блокує роботу гена. Якщо ділянка вбудовування знаходиться поряд з геном, то регуляторні елементи вірусу можуть впливати на роботу клітинних генів. Вбудовування «чужих» регуляторних послідовностей поряд з генами, що відповідають за чергування фаз ділення і зростання клітки, приводить до переродження клітки в ракову. При вбудовуванні генома вірусу в зародкову лінію кліток вірусна ДНК успадковується як менделирующий ознака. Більшість вірусних послідовностей вбудувалися в геном предків людини десятки мільйонів років назад. За минулий час в них накопичилася безліч мутацій і вони втратили свою патогенність. Частина з них зберегла здатність «стрибати» по геному, переносячи регуляторні елементи. Ендогенні ретровіруси складають близько 3% ДНК людини. Гени людини (також як і інших эукариотических організмів) мають складну структуру. Після синтезу РНК деякі її ділянки (їх називають вставними послідовностями або интронами) вирізуються, а ті, що залишилися (їх називають экзонами) зшиваються в єдиний ланцюг, белок-кодирующую послідовність, що містить, і сигнали регуляції трансляції. Інтрон-екзонная структура генів достатньо складна. Екзони можуть з'єднуватися в різних поєднаннях, завдяки чому один ген може визначати синтез декількох десятків що розрізняються своєю амінокислотною послідовністю білків. Відмінності интрон-экзонной структури зрілої мРНК можуть визначати інтенсивність синтезу одного і того ж білка в різних тканинах або на різних етапах онтогенезу. Більшість генів в кожній клітці «мовчать». Набір активних генів розрізняється залежно від типу тканини, періоду розвитку організму, отриманих зовнішніх або внутрішніх сигналів. Можна сказати, що в кожній клітці «звучить» свій акорд генів, визначаючи спектр мРНК, що синтезуються, кодованих ними білків і, відповідно, властивості клітки. У кожній клітці (окрім еритроцитів, у яких відсутнє ядро) працюють гени, що кодують ферменти реплікації і репарації ДНК, транскрипції, компоненти апарату трансляції (рибосомные білки, рРНК, тРНК, аминоацилсинтетазы і ін. ферменти), ферменти синтезу АТФ і інші компоненти, необхідні для ведення «домашнього господарства» клітки. Завідують «домашнім господарством» біля однієї п'ятої всіх генів. ДНК-рівень. Загальна кількість ДНК в соматичній клітці складає 6,4 х109 пару нуклеотидов, отже, гаплоидный набір складається з 3,2х109 пар нуклеотидов. Основна кількість ДНК локалізована в хромосомах (95%). Внехромосомная частина генома людини — ДНК мітохондрій (5%). Зовсім невелику кількість складають окремі кільцеві молекули ДНК в ядрі і цитоплазмі. По способах організації нуклеотидов і функцій сегментів ДНК можна виділити наступні фрагменти (рис.29): 1) гени мРНК (структурні гени); 2) гени рРНК; 3) гени тРНК; 4) ДНК сателіта; 5) спейсерная ДНК.
Ріс.29. Структурна організація нуклеотидных послідовностей (генів) в ДНК
Структурні гени (тисячі різновидів) несуть інформацію про структуру певних поліпептидів. З цих ділянок ДНК транскрибируется мРНК, яка направляє синтез білків. Гени рРНК (декілька різновидів) містять інформацію про структуру рибосомальных РНК і обуславливают їх синтез. Гени тРНК (більше 30 різновидів) несуть інформацію про транспортні РНК. ДНК сателіта представлена великим числом груп нуклеотидов, що повторюються, в різних ділянках ДНК, яка не має сенсу і не транскрибируются. Значення ДНК сателіта повністю поки не відоме. Спейсерная ДНК розділяє між собою гени, вона не транскрибируется. Роль цих ділянок до кінця не з'ясована. Різні структурні гени мають особливості організації (рис.29 Би). Наприклад, гени, що повторюються, – один і той же ген багато разів повторюється (багато сотень разів), не відділяючись один від одного, утворюючи тандеми (наприклад, гени рРНК). Кластери генів – це групи різних структурних генів на певній ділянці хромосоми, об'єднаних загальними функціями. Наприклад, кластери п'яти разных гистонов повторюються по 10-20 разів. Одиночні гени серед ДНК сателіта зазвичай мають регуляторну або підсилюючу дію на структурні гени, наприклад, энхансеры. Внехромосомниє і кільцеві молекули ДНК виявляються в цитоплазмі і ядрі. У людини вони вивчені ще недостатньо. У строгому сенсі вони є не складовими елементами генома, а його продуктом. Їх розмір коливається від 150 до 20 000 пар нуклеотидов. Є ці молекули продуктом фрагментації хромосомної ДНК в клітці або утворюються за рахунок інших генетичних процесів (гомологичная рекомбінація, зворотна транскрипція), поки неясно. Досліджені до теперішнього часу у ссавців великі кільцеві молекули ДНК розміром від 150 до 900 000 пар нуклеотидов, локалізовані тільки в ядрах, є амплифицированные ділянки онкогенов або генів стійкості до отрут і антиметаболитам. З цими молекулами імовірно зв'язують стійкість кліток до ліків і здатність кліток до необмеженого зростання. Їх походження пояснюють делециями відповідних областей хромосом. Хромосомна ДНК підрозділяється на дві групи ділянок: з унікальною послідовністю пар нуклеотидов і з послідовностями, що повторюються. Із загальної маси ДНК в клітці приблизно 50% ДНК з унікальними послідовностями і 50% — з тими, що повторюються. Частина ДНК, що кодує білки складає всього 3—5%. Що робить частина генома, що «покоїться», невідомо. Проте важко припустити, що вона не має функцій. Поліморфізм. Будь-які зміни в структурі ДНК (у хромосомах або мітохондріях) ведуть до генетичного поліморфізму. Ці зміни можуть бути якісними, якщо вони обумовлені заміною або втратою нуклеотидов, або кількісними, якщо в певному локусе варіює число нуклеотидных повторів різної протяжності. І ті та інші варіанти генетичного поліморфізму зустрічаються як в смислових (внутриэкзонных), так і в несмислових (внегенных або интронных) послідовностях молекули ДНК. Головною формою генетичного поліморфізму є однонуклеотидный поліморфізм (ОНП). Під цим терміном розуміють варіанти послідовностей ДНК у різних людей із залученням однієї пари нуклеотидов (рис.30).
Ріс.30. Приклади нуклеотидного поліморфізму у двох індивідів
На даному малюнку представлено три фрагменти послідовностей від двох індивідів. У прямокутниках виділені однонуклеотидные відмінності в послідовностях геномів. ОНП — найбільш загальне джерело варіацій між людьми. Ці варіації зустрічаються впродовж всієї ДНК (у экзонах, интронах, міжгенних проміжках, повторах) і відображають минулі мутації. Секвенірованієм геномів або їх частин різних людей встановлено, що однонуклеотидные відмінності виявляються впродовж 1000—2000 нуклеотидной довжини. Це означає, що на всю довжину генома (3,2 млрд пар нуклеотидов) повинне бути 1,6—3,2 млн ОНП. До 2001 р. ідентифіковано і картировано 1,42 млн ОНП. Розрахунки показують, що дві люди на 99,9% ідентичні по нуклеотидным послідовностях, тобто тільки 0,1% відмінностей по одному нуклеотиду створює такі величезні індивідуальні фенотипічні варіації, які легко бачити в будь-якій групі індивідів. Припускають, що відмінності по одній підставі між певними відрізками геномів лежать не тільки в основі генних хвороб (миссенс-мутации), але і в основі чутливості до збудників або захисту від них, в основі пристосовних реакцій і спадкового нахилу до мультифакториальным хвороб. На початок 2001 р. ідентифіковане 60 000 ОНП в генах (їх називають кодуючими ОНП). Це означає, що в генних послідовностях один кодуючий ОНП зустрічається в межах 1080 пар нуклеотидов. Хоча інформація про ОНП ще не повна (основні відомості отримані в останніх 2 року), вже відомо, що 93% генів містять ОНП. Головне використання карти ОНП — з'ясування внеску індивідуальних генів в хворобі комплексної (багаточинником) і полігенної природи. Порівняння частот певних типів ОНП у пацієнтів і в контрольних групах дозволяє ідентифікувати ОНП, з якими асоціюється захворювання. Не дивлячись на великі перспективи, які відкриваються для пояснення захворювань людини з розумінням природи і розмаху ОНП, необхідно пам'ятати про небезпеку геномомании. Гени і геноми діють не у вакуумі. Середовище не менш важливе для біології людини, чим гени. Карти ОНП при правильному використанні дозволяють краще зрозуміти роль природи (генотипу) і середовища в широкому розумінні в розвитку людини в цілому і патології зокрема. Генетичні карти. Складовою частиною відомостей про геном людини разом з нуклеотидной послідовністю є генетичні карти хромосом, тобто схеми, що описують порядок розташування генів і інших генетичних елементів на хромосомі з вказівкою відстані між ними. Генетична відстань вимірюється по частоті рекомбінації між гомологичными хромосомами і виражається в сантиморганидах (сМ). Одна сМ відповідає частоті рекомбінації, рівної 1%. Довжина всього генома людини рівна приблизно 3000-3500 сМ.
Ріс.31. Патологічна анатомія хромосоми 3
Знання генетичних карт необхідне в різних розділах медичної генетики: для діагностики хвороб методом зчеплення; оцінки патологічних ефектів хромосомних транслокаций; вирішення питань еволюційної і популяції генетики. Гени, що кодують поліпептиди, РНК. Гени, що кодують білки, як правило, містять кодуючі області (экзоны), що перериваються одним або більш нитронами. Проте деякі гени (наприклад, гистоновые або кодуючі інтерферони) не мають вставних послідовностей. Число таких послідовностей сильно варіює від гена до гена, а кількість ДНК, що припадає на частку интронов, у багато разів перевищує кількість ДНК кодуючих областей — у деяких організмів в десятки разів. Як правило, нуклеотидные послідовності аналогічних экзонов, що відносяться до паралогичным генів в даному геномі або до ортологичным генів в геномі різних видів, консервативніші, ніж нуклеотидные послідовності відповідних интронов. Положення интронов, як правило, фіксоване, а по довжині і складу вони варіюють. Всі відомі гени, що кодують білки, транскрибируются РНК-полімеразой II і тому часто мають схожі промоторы і сигнали полиаденилирования. Але багато такі гени пов'язані з більш специфічними регуляторними послідовностями, які опосередкують дію гормональних, средовых або онтогенетичних чинників. На відміну від генів, кодуючих РНК, полипептидные гени представлені в геномі в однині, проте при цьому геном часто містить сегменти, гомологичные даному специфічному гену. Таким чином, однокопийный ген може входити до складу сімейства близькоспоріднених послідовностей (наприклад, в сімейство генів гормону зростання). Члени такого сімейства можуть кодувати білки, що трохи розрізняються (наприклад, изозимы). Проте вони можуть мати різні регуляторні сигнали, відповідальні за експресію генів в різних тканинах або на різних стадіях розвитку (як у разі генів гормону зростання або плацентарного лактогена). Рівним чином членами сімейства можуть бути і псевдогени. Умовилися, що два неаллельных гена вважаються ідентичними, якщо вони кодують фактично однакові білки і знаходяться під загальним контролем. Мультігенниє сімейства. Сімейства генів зазвичай кодують білки, багато представлені в клітці. У своїй більшості гени активні. По критерію відповідності послідовностей специфічному зонду виявляються багато генів тих, що кодують синтез інтерферону, актина, тубулина... Усередині сімейства генів деякі його члени можуть повторюватися частіше, ніж інші. Наприклад: існує 8 генів для а-типа інтерферону людини, і лише один в-типа. Багато генів можуть бути об'єднані в кластери або розсіяні по геному. Загальний план будови глобиновых генів консервативний. Гени-інтерферони мають схожу структуру, для якої характерна відсутність интронов. Гени актина мають переривисту структуру. У цих генів ділянки, що кодують білок, володіють високим ступенем гомологии. Таким чином, якщо активні гени відбулися від загального гена-предка, розташування экзонов і интронов зазнало істотні зміни. Функціонуючі гени можуть мати переривисту і безперервну будову і зміни в розташуванні интронов не обов'язково впливають на активність генів. Гени, що кодують однакові або близькі білки, необов'язково повинні бути організовані у вигляді тандему, але можуть бути розсіяні по геному у вигляді окремих індивідуальних генів або малих кластерів. Повторення послідовностей приводить до збільшення кількості ДНК, що кодує певну функцію. Можливе існування декількох активних генів, що кодують або один і той же білок, або несподівані його варіанти. Коли копії генів об'єднані в кластери, відстань між ними може бути значною, що може збільшувати загальну кількість ДНК, відповідальну за здійснення даної функції: основна частина функції кодується декількома генами, а не унікальним геном; значна кількість ДНК не несе кодуючої функції. Однокопійниє гени можуть також належати великому сімейству віддалено споріднених послідовностей. Не дивлячись на структурну схожість, гени такого надсемейства кодують абсолютно різні білки (як, наприклад, вже згадані гени пролактина і гормону зростання). Розподіл окремих генів по надсемействам не завжди вдається провести цілком чітко. Часто схожість нуклеотидных послідовностей членів варіює від 95% і більш до 50% і менш. У жорстких умовах відпалу дуплекси утворюють тільки сегменти ДНК з гомологией не менше 80%, тому неблизькоспоріднені сімейства можна порівнювати, лише знаючи їх нуклеотидную послідовність. Більш того, схожість між двома білками не знаходиться в простій залежності від процентної схожості між відповідними генами. Унаслідок вырожденности генетичного коду значні зміни в третій позиції кодонів мало впливають на тип кодованої амінокислоти, а такі ж зміни в першій або в другій позиціях приводять до істотних змін в структурі білка. Крім генів, що належать мультигенным сімействам або надсемействам, є і унікальні гени (наприклад, ген тиреоглобуліну людини і ген актина дріжджів). Кожному мультигенному сімейству генів, що кодують білки, властиві особливі властивості, характерні для даного вигляду. Актіновиє гени: мулътигенное консервативне сімейство. Актин бере участь в самих різних типах клітинного руху, зокрема він забезпечує клітинну рухливість і м'язові скорочення. Відповідно эукариоты мають декілька різних типів актина і генів, що кодують їх. Одні з цих генів організовані в кластери, інші диспергированы. В ході еволюції актиновых генів в них вбудовувалися нові интроны. Кожне така подія приводила б до зсуву сусідніх послідовностей і до зрушення рамки прочитування. Можна припустити зворотне: більшість актиновых генів містили интроны у всіх тих положеннях, де вони знаходяться і в сучасних генах, але при еволюції разных таксонов втрачалися різні интроны. Актіновиє гени в основному розсіяні по хромосомах. У миші, наприклад, гени а-актина скелетних м'язів і серцевого м'яза локалізовані в хромосомах 3 і 17 відповідно, а гени цитоплазми в-актина ~ у хромосомі 5. У більшості вивчених організмів число актиновых генів, мабуть, перевищує число відомих актиновых білків. Хоча не виключено, що якісь з цих «экстра»- послідовностей кодують не виявлені поки форми актина, більшість з них, ймовірно, є псевдогенами, зокрема процессированными. У людини щонайменше дві з двадцяти актиноподобных послідовностей є процессированные псевдогенами в-актина; вони не містять интронов і несуть в кодуючих областях різні мутації. Тубуліновиє гени: мультигенное сімейство, яке включає гени два разных субъединиц гетеродимерного білка. Мікротрубочки беруть участь в багатьох процесах, що протікають у всіх эукариотических клітках: мейозе, митозе, клітинному русі і секреції. Тому не дивно, що структура тубулина-білка, з якого складаються мікротрубочки, так само як і нуклеотидная послідовність гена, що кодує тубулин, однакові у всіх эукариот. кДНК, синтезовані на тубулиновой мРНК курки, гибридизуются з тубулиновыми генами таких віддалених організмів, як дріжджі і ссавці. Тубулін - це гетеродимер, що складається з двох поліпептидів: а і ст. а- і в-субъединицы містять 450-451 і 445 амінокислотних залишку відповідно і гомологичны приблизно на 40%. Гени обидві субъединиц відносяться до одного мультигенному суперсімейства, хоча досить сильно розрізняються і не гибридизуются. Як правило, сімейства тубулиновых генів у кожного виду эукариот кодують разные изотипы а- і в-субъединиц. Амінокислотні послідовності різних а- або в-субъединиц, що кодуються цими генами, зазвичай розрізняються лише трохи (менш ніж на 10%), і відмінності стосуються в основному карбоксильних кінців молекул. Мабуть, варіації в структурі тубулинов пов'язані з тим, що трохи розрізняються ті мікротрубочки, які вони утворюють. Останні у свою чергу специфічні відносно клітинних процесів, типів кліток і стадій розвитку. Таким чином, разные а- і в-изотипы можуть відповідати різним типам мікротрубочок, що виконують певні функції, хоча така спеціалізація і не абсолютна. Подальші зміни в структурі тубулина відбуваються вже після транскрипції і теж сприяють функціональній спеціалізації мікротрубочок. Про таку спеціалізацію свідчить висока консервативність специфічних изотипов у хребетних. Наприклад, у різних хребетних переважаючий в нервовій тканині в-тубулин має на карбоксильному кінці абсолютно однакові послідовності. Суперсімейства генів і їх продукти. Суперсімейство глобиновых генів. Гемоглобін хребетних — це гетеротетрамеры, що містять по два а- і в-полипептида. У геномі всіх хребетних міститься безліч генів і псевдогенів а- і в-цепей- членів суперсімейства, яке включає кодуючі послідовності генів глобінів безхребетних, міоглобіну хребетних і леггемоглобина рослин. Всі ці білки містять гем і оборотно зв'язуються з киснем. Глобіновиє мРНК і відповідні гени були серед перших об'єктів, які досліджувалися методами рекомбинантных ДНК. Гемоглобіну припадає на частку більше 90% всіх розчинних білків еритроцитів, а на долю глобиновой мРНК — велика частина мРНК ретикулоцитов і ядерних еритроцитов. Ні у яких інших клітках глобиновые гени в помітній кількості не транскрибируются. Можливість отримання щодо чистої глобиновой мРНК, наявність величезного числа вже відомих мутацій в генах глобіну людини, а також обширні дані про властивості глобиновых білків послужили стимулом до молекулярних досліджень цих генів. Вдалося отримати перші дані про интронах в клітинних генах і про властивості промоторов для РНК-полімерази II. Була встановлена нуклеотидная послідовність різних аллелей глобиновых генів у людини і інших видів, що послужило основою для створення численних теорій еволюції глобінів. Як правило, у ссавців є безліч глобиновых генів і псевдогенів. У людини в-глобиновые гени об'єднані в кластер завдовжки 65 т.п. н., розташований в хромосомі II, а а-глобиновые гени — в кластер завдовжки 25 т. п.н. у хромосомі 16. Кластер в-глобиновых генів містить п'ять генів, а e-глобиновый кластер — три. Обидва кластери включають також по декілька псевдогенів. У людини і інших ссавців різні глобиновые гени экспрессируются на різних стадіях розвитку організму. У багатьох видів, у тому числі і у людини, розташування генів в а- і в-кластерах відповідає тому порядку, в якому вони экспрессируются під час розвитку організму. Інтерфероновиє гени. У відповідь на різноманітні зовнішні дії клітки багатьох хребетних секретируют поліпептиди, звані інтерферонами. Наприклад, в результаті вірусної інфекції і попадання в клітку двухцепочечной РНК в лейкоцитах індукується синтез інтерферонів групи e, а у фибробластах-интерферонов групи в (IFN-а, I IFN-в, 1 IFN-г). Кожна група містить різні структурні споріднені білки. Абсолютно інший білок, унікальний lFM-y, синтезується в лімфоцитах при реплікації ДНК, індукованою митогенами. Всі ці інтерферони у свою чергу викликають різні клітинні ответы- придушення вірусної інфекції, імунні реакції, протипухлинну активність. Потенційна значущість інтерферонів як терапевтичних засобів для лікування вірусних інфекцій і ракових захворювань стимулювала їх дослідження. Серед клонованих сегментів ДНК людини, що містить lFN-a-гены, деякі несли більш за один IFN-a-гена. Ці дані, а також виявлення методом гібридизації in situ єдиного IFN-a-локуса в геномі людини говорять про те, що гени, що кодують інтерферони групи, утворюють одиничний кластер. Генетичний аналіз показує, що найближчими сусідами генів IPN-a на хромосомі 9 є гени IPN-fi. Єдиний ген IFN-y розташований на іншій хромосомі. Сусідні гени схожі по своїй структурі. Псевдогени. Їх називають так тому, що вони містять нуклеотидные послідовності, схожі з послідовностями функционально- активних генів, але не можуть экспрессировать з утворенням фунционально - активного білка. Деякі псевдогени мають в цілому таку ж структуру, як і функціонально-активні гени, із звичайним розташуванням послідовностей відповідних экзонам і интронам. Вони становяться не активними в результаті мутації, що порушують одну або всі стадії експресії гена. Ці зміни можуть виявлятися у вигляді ініціації транскрипції, перешкоджати здійсненню сплайсинга на межах экзон-интрон або приводити до того, що передчасному термінує трансляції. Зазвичай псевдоген несе декілька тимчасових мутацій, тому що ген одного разу переставши бути активним став об'єктом для подальшого накопичення мутацій. Такі псевдогени виявлені в багатьох системах генів, включаючи гени глобінів, иммуноглобинов, антигенів гистосовместимости і так далі Наприклад: псевдоген кролика із звичайною організацією экзонов і интронов по будові близький до функционально- активному гену, але в кодоні 20 псевдогена є делеция однієї пари нуклеотидных підстав, що викликає зрушення рамки прочитування, із-за якого трансляція термінує незабаром після її початку. В результаті точковых мутацій виявилися зміненими декілька розташованих правіше кодонів, що кодують амінокислоти, наявні у всіх глобінах, але один з 2-х интронов псевдогена не зберіг прикордонні послідовності, що задовольняють правилу. Тому интроны не можуть бути видалені при сплайсинге навіть якщо б ген і транскибировался. Загальний вивід, виходячи із структури таких псевдогенів - це незалежний характер еволюції кожного з них в процесі еволюції кластера глобіну генів кожного виду організмів. Таким чином, виникнення нових генів, за якими слідує їх закріплення в геномі як функціональні копії, їх зміна приводить до утворення нових функціонально-активних генів, або инактивация з утворенням псевдогенів - процеси що відбуваються в кластері постійно. Псевдогени, які мають схожість з Рнк-транськріптом, називаються процессироваными псевдогенами. Псевдогени являються членами більшої частини сімейства генів. Зазвичай псевдогени складають дуже невелику частину від загального числа генів. Але є виключення: рибосомный білок миші кодується одним активним геном, що має близько 15 схожих з ним процессированных псевдогенів. Псевдогени- це безвихідь еволюції, просто небажаний побічний ефект перебудов функционально- активних генів. Механізми, відповідальні за дупликации, делеции і перебудови генів, впливають на всі послідовності, що відносяться до членів кластера, незалежно від того, є вони функціонально - активними чи ні. Область, що включає псевдогени, - це повтор послідовностей, що оточує початок активного гена (від -73 до +101 п.н.). Дивовижна особливість кластера в цілому полягає в тому, що він, таким чином, повинен містити однакове число генів і псевдогенів. Наприклад, промотор транскрипції псевдогена 5S РНК цілком розташований усередині гена. Цей псевдоген преобрел відмінності від гена в результаті всього лише дев'яти точковых мутацій. Таким чином, до складу псевдогена входить ділянка, що містить злегка змінений внутрішній промотор гена. Необхідне розобраться є ці зміни достатніми для того, що б псевдоген втратив здібність до транскрипції з утворенням РНК - продукту in vivo. У клітці могли проходити РНК залежні транспозиції, що приводять до утворення псевдогенів. Деякі псевдогени - проявляють такі зовнішні і внутрішні ознаки які свідчать про можливість їх походження з послідовності РНК. Початок псевдогена відповідає крапці еквівалентною 5'-концу РНК, і саме це свідчить про походження його ДНК з РНК. Декілька псевдогенів складається із зчленованих послідовностей экзонов, що доводить посередництво РНК при утворенні псевдогена. Псевдоген може закінчуватися короткою областю з А-Т пар підстав. Від обох його сторін є короткі прямі повтори. Якщо псевдоген відбувся з послідовності м-РНК його гомология в 5'-конце не може розповсюджуватися на послідовності розташовані вище за сайт ініціації. Транспозони. Потенційна можливість для зміни прокариотических і эукариотических геномів забезпечується здатністю певних послідовностей переміщатися з одного сайту в іншій - ці послідовності отримали назву транспозонов. Транспозиція не заснована на якій-небудь спорідненості між послідовностями в донорных і реципиентных сайтах, і це відрізняє його від інших механізмів, що беруть участь в перебудовах ДНК. Кожен транспозон несе гени, потрібні для його власної транспозиції, хоча для цього необхідні і функції генома, в яких знаходяться транспозоны. Транспозони можуть створювати в клітинних системах часткові області гомологии, оскільки їх копії в різних місцях (різні хромосоми) забезпечують можливість реципрокной рекомбінації. Такі обміни можуть приводити до делеции, инсерциям, інверсіям або транслокациям. Транспозиційні механізми беруть участь в цілому ряду подій, від з'єднання негомологичных послідовностей ДНК до забезпечення специфічних рекомбінаційних процесів. Будь-яка транспозиційна подія може забезпечити селективні переваги. Наприклад: генетичні перебудови обуславливает переважну виживає генома, що несе активний транспозон. Транспозірующиеся елементи були відкриті при виявленні вставок (инсерций) нового матеріалу в межах бактерійних оперонів. Транспозиція пов'язана з утворенням додаткової копії транспозона в реципиентном сайті. Більшість транспозонов мають декілька сайтів впровадження. Транспозон - автономна одиниця, яка кодує білки, необхідні для транспозиції. Реакція включає пізнавання кінців транспозирующегося елементу. Найкоротші транспозоны кодують тільки білки, учавствующие в транспозиції. Крупніші транспозоны несуть додаткові генетичні маркери. Транспозони мають регуляторні сигнали, що відносяться до їх власних генів, ці сигнали іноді можуть впливати на події в оперонах. У деяких транспозонах поблизу їх меж містяться промоторы, в яких ініціюється транскрипція фланкирующего матеріалу: активуються при цьому гени, суміжні з елементом. Транспозони мають инвентированные кінцеві повтори. Впровадження транспозонов викликає освіта в сайті-мішені прямих повторів фланкирующей ДНК. Інсерционниє послідовності - це прості транспозоны. Перша група виділених транспозонов отримала назву инсерционных послідовностей. IS-элемент є простий клас транспозонов, їх генетичні функції пов'язані тільки із здібністю до транспозиції не дивлячись на те, що кожен IS-элемент має властиву тільки йому послідовність. Форма організації у них загальна. Кожен елемент володіє короткими інвертованими кінцеви
|
|||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-09-13; просмотров: 773; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.96 (0.017 с.) |

 Вивчення груп зчеплення і складання карт хромосом спочатку грунтувалися на аналізі «розщеплювання» фенотипов в потомстві формально-генетичними методами. Застосування молекулярно-генетичних методів значно прискорило картирование генів, а секвенирование генома дозволяє скласти повні генетичні карти для всіх хромосом. На рис.31 представлена як приклад карта хромосоми 3 по генах, патологічні мутації в яких ведуть до спадкових хвороб. Такі карти називають патологічною анатомією генома людини. Слід зазначити, що це далеко не повна карта, вона постійно уточнюється.
Вивчення груп зчеплення і складання карт хромосом спочатку грунтувалися на аналізі «розщеплювання» фенотипов в потомстві формально-генетичними методами. Застосування молекулярно-генетичних методів значно прискорило картирование генів, а секвенирование генома дозволяє скласти повні генетичні карти для всіх хромосом. На рис.31 представлена як приклад карта хромосоми 3 по генах, патологічні мутації в яких ведуть до спадкових хвороб. Такі карти називають патологічною анатомією генома людини. Слід зазначити, що це далеко не повна карта, вона постійно уточнюється.



