
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Провідникова функція спинного мозку. Залежність спінальних рефлексів від діяльності центрів головного мозку . Спінальний шок.Содержание книги
Поиск на нашем сайте
Біла речовина спинного мозку (передні, бокові та задні канатики) складається з нервових волокон, які формують провідні шляхи. Їх поділяють на висхідні та низхідні. Основними висхідними шляхами є: 1. Шлях Голя – розташований в медіальній частині заднього канатика. 2. Шлях Бурдаха – розташований в латеральній частині заднього канатика. Ці два шляхи є аферентними шляхами, що проводять пропріоцептивну чутливість кіркового напрямку (усвідомлену). 3. Задній спинномозково-мозочковий шлях – задня частина бокового канатика. 4. Передній спинномозково-мозочковий шлях – розташований вентральніше попереднього. Ці два шляхи передають інформацію від пропріорецепторів в мозочок. 5. Спіноталамічні шляхи – розташовуються в бокових канатиках вентрально та попереду від попередніх. Передають тактильну, больову, температурну чутливість. Основними нисхідними шляхами є: 1. Бічний кірково-спинномозковий шлях (пірамідний) проводить імпульси свідомих рухів – бокові канатики. 2. Червоноядерно-спинномозковий шлях (екстрапірамідний) проводить імпульси несвідомих рухів та тонусу м’язів – бокові канатики. 3. Передній кірково-спинномозковий шлях (пірамідний) проводить імпульси свідомих рухів – передні канатики. 4. Присінково-спинномозковий шлях – передні канатики. 5. Сітчасто-спинномозковий шлях – передні канатики. Ці два шляхи проводять імпульси несвідомих рухових реакцій, які спрямовані на збереження рівноваги тіла та координацію рухів. Низхідні шляхи спинного мозку об’єднують в дві системи регуляції рухових функцій – латеральну та медіальну. 1. Латеральна низхідна система (ЛНС) регуляції рухових функцій.
2. Медіальна низхідна система (МНС) регуляції рухових функцій.
Залежність діяльності спинного мозку від низхідних впливів вищих рівнів ЦНС проявляється, якщо ці впливи припинити (шляхом перерізки між спинним та заднім мозком, чи шляхом охолодження в цій ділянці). В результаті припинення впливів вищих відділів ЦНС на спинний мозок розвивається спінальний шок – тимчасове припинення рефлекторної діяльності спинного мозку. У жаби явище спінального шоку продовжується декілька хвилин, у кішки – декілька днів, у приматів – декілька неділь. У приматів після відновлення рефлекторної діяльності спинного мозку частина рефлексів змінюється в порівнянні з інтактними тваринами, а частина не відновлюється зовсім. Ведучим механізмом в розвитку спінального шоку є припинення активуючих впливів структур головного мозку на нейрони спинного мозку. Чим вищий рівень організації тварини, тим більш важливими є ці впливи. 24. Рухові рефлекси заднього мозку, децеребраційна ригідність. Задній мозок – це довгастий мозок та міст. Вони зберігають ознаки сегментарної будови, але мають і надсегментарні утворення. Серед найбільш важливих рухових надсегментарних утворень – присінкові ядра та сітчаста речовина. Вони носять назву надсегментарних утворень, так як впливають на м’язи не прямо, а через мотонейрони сегментарних структур – рухові ядра спинного мозку і черепно-мозкових нервів. Задній мозок отримує і переробляє всю аферентну інформацію, що надходить від спинного мозку, оскільки всі специфічні висхідні шляхи від спинного мозку, входячи в стовбур мозку (задній та середній мозок) віддають коллатералі (гілочки) до ретикулярної формації Ù тут продовжується обробка аферентної інформації. Задній мозок отримує нову аферентну інформацію, яка відіграє роль в регуляції рухових функцій – це інформація від вестибулярних рецепторів.
Отолітові рецептори – являють собою волоскові клітини, волоски яких занурені в отолітову мембрану – желеподібна мембрана з включенням кристалів CaCО3 – отолітів. Зміщення цієї мембрани деформує волоски Ù збудження рецептора (чи його гальмування, в залежності від того, в який бік згинається волосок). Отолітова мембрана зміщується відносно волоскових клітин при двох умовах: 1. Зміщення голови в просторі. 2. Рух з лінійним прискоренням. Вестибулорецептори півколових канальців – волоскові клітини, які локалізуються в ампулах півколових канальців і занурені в ендолімфу цих канальців. Ендолімфа зміщується відносно волоскових клітин і деформує їх при русі з кутовим прискоренням. При цьому збуджуються волоскові клітини канала, який знаходиться в тій площині, в якій проходять обертальні рухи (у відповідності з трьохвимірним простором, в якому ми існуємо, наявні 3 півколових канальці). Від вестибулорецепторів інформація передається в задній мозок по аферентним волокнам VIII пари черепно-мозкових нервів (далі ЧМН) – вестибуло-кохлеарного нерва (точніше – по його вестибулярній частині). В задньому мозку розміщені 4 вестибулярні ядра (медіальне, латеральне, верхнє та нижнє). В регуляції рухових функцій у людини найбільше значення має латеральне вестибулярне ядро Дейтерса, від якого починається вестибулоспінальний шлях (частина МНС). Вивчити роль заднього мозку в регуляції рухових функцій можна на тваринах з децеребрацією – перерізкою стовбура мозку між середнім та заднім мозком (точніше – нижче червоних ядер середнього мозку). При порівнянні можливостей регуляції рухових функцій такої тварини і спінальної тварини, можна оцінити роль в регуляції цих функцій саме заднього мозку.
Активація a-мотонейронів екстензорів приводить до безпосередньої активації екстрафузальних волокон цих м’язів Ù збільшення їх тонуса. Активація g-мотонейронів приводить до скорочення скоротливих сегментів інтрафузальних волокон Ù скорочення цих сегментів Ù розтягнення ядерної сумки інтрафузального волокна Ù збудження рецептора Ù передача інформації по g-петлі до a-мотонейронів Ù посилення їх активності Ù збільшення тонусу м’язів екстензорів. Протилежні зміни виникають у флексорній групі м’язів. Тому коротко можна сказати, що в основі розвитку децеребраційної ригідності лежать вестибулоспінальні впливи, завдяки яким відбувається: - a- та g-посилення міотатичних тонічних рефлексів екстензорів; - a- та g-послаблення міотатичних тонічних рефлексів флексорів. Шийні рефлекси положення – виникають у відповідь на зміну нормального положення голови відносно тулуба. При цьому подразнюються пропріорецептори шиї Ù передача інформації в шийні сегменти спинного мозку Ù по пропріоспінальним низхідним шляхам до мотонейронів спинного мозку Ù перерозподіл м’язового тонусу, який змінює положення тулуба відповідно до положення голови: - якщо голова нахиляється чи повертається в бік, то зі сторони нахилу збільшується тонус розгиначів, а з протилежної сторони – тонус згиначів. Такий перерозподіл тонуса попереджає падіння, тобто втрату пози; - якщо голова нахиляється вперед, тонус згиначів збільшується на передніх кінцівках кішки, тонус розгиначів – на задніх (кицька п’є молоко чи жере Віскас); - якщо голова закидається назад, тонус розгиначів збільшується на передніх кінцівках кішки, тонус згиначів – на задніх (кицька дивиться на папугу й готується до стрибка). Присінкові (вестибулярні) рефлекси положення – виникають при зміні положення голови в просторі. При нахилах чи поворотах голови збуджуються отолітові вестиблорецептори Ù інформація передається до вестибулярних ядер Дейтерса Ù до мотонейронів спинного мозку та до м’язів. В результаті зі сторони нахилу (поворота) збільшується тонус розгиначів, а з протилежної сторони – тонус згиначів Ù збереження пози.
Перерозподіл тонусу за рахунок шийних рефлексів не можна отримати у спінальної жаби через слабкість тонуса, хоча рівень їх замикання – спинний мозок. Задній мозок підсилює ці рефлекси. Шийні та присінкові рефлекси положення у вигляді перерозподілу тонуса можна отримати у децереброваної тварини. Але на фоні ригідності ступінь цього перерозподілу недостатній для утримання антигравітаційної пози: децеребрована тварина може стояти, як лялька, але ще не може втримувати рівновагу при умовах, що вмикають шийні та присінкові статичні рефлекси положення. Таким чином, задній мозок: 1. Забезпечує створення посиленої антигравітаційної пози – пози децеребраційної ригідності. 2. Забезпечує здійснення статичних позових рефлексів (шийних та присінкових), що спрямовані на збереження рівноважної пози при зміні положення голови відносно тулуба (шийні) чи при зміні положення голови в просторі (присінкові), але ступінь виразності цих рефлексів на фоні посиленого тонуса недостатній для збереження пози. 3. Не забезпечує збереження пози при русі (немає стато-кінетичних рефлексів), відновлення порушеної пози (немає випрямляючих рефлексів). 4. Фазичні рефлекси на фоні посиленого тонуса не викликаються. Ретикулярна формація стовбура мозку, характер її висхідних та низхідних впливів. Сітчаста речовина (ретикулярна формація – РФ) – скупчення нервових клітин, які мають велику кількість дендритів, що галузяться та довгі аксони, які формують низхідні (ретикулоспінальні) та висхідні шляхи сітчастої речовини. Аферентних шляхів РФ дуже багато, вона отримує інформацію від: - усіх специфічних висхідних еферентних шляхів, які віддають колатералі до ядер РФ, заходячи в стовбур мозка; - від іних утворень ЦНС, а саме: - від кори головного мозку; - від гіпоталамуса; - від мозочка; - від базальних ядер; - від інших утворень. Еферентні зв’язки РФ можна поділити на висхідні та низхідні. Висхідні зв’язки РФ та висхідні впливи: - через неспецифічні ядра таламуса передає інформацію всім відділам головного мозку та має неспецифічний активізуючий вплив на клітини. Цей вплив забезпечує бодьорий стан організму та визначає рівень активності клітин кори. РФ має окремі структури (гіпногенні зони), котрі зменшують ступінь (рівень) активності клітин кори та зумовлюють стан сну; - по ретикулогіпоталамічним шляхам передає інформацію від різних рецепторів в гіпоталамус. Характер низхідних ретикулоспінальних впливів: - від медіального ядра РФ заднього мозку починається ретикулоспінальний (входить до складу МНС) – специфічно активує мотонейрони флексорів та гальмує мотонейрони екстензорів тулуба та проксимальних відділів кінцівок; - від інших ядер РФ беруть початок шляхи, які неспецифічно активують чи гальмують спінальні мотонейрони і таким чином визначають ступінь їхньої активності; - від вегетативних центрів РФ (дихальний, гемодинамічний) по ретикулоспінальним шляхам передається інформація до симпатичних рухових нейронів спинного мозку при регуляції відповідних функцій організму. 25. Рухові рефлекси середнього мозку, їх фізіологічне значення. Середній мозок (СрМ) за участі сітчастої речовини опрацьовує аферентну інформацію, яка поступає в спинний та задній мозок. Нова інформація поступає в СрМ від зорових та слухових рецепторів. Її опрацювання відбувається в ядрах передніх (від зорових рецепторів) та нижніх (від слухових рецепторів) горбків. На основі опрацьовання інформації від усіх цих рецепторів СрМ здійснює контроль за станом зовнішнього та внутрішнього середовища організма. Важливими надсегментарними руховими ядрами СрМ є: 1) червоні ядра – від них інформація від нейронів спинного мозку передається по шляхах що перехрещуються (руброспінальні шляхи – елемент ЛНС); 2) ретикулярна формація; 3) чорна субстанція – знаходиться в середньому мозку, але функціонує як єдине ціле з базальними ядрами. Руброспінальні впливи характеризуються активацією a- та g-мотонейронів флексорів та гальмуванням a- та g-мотонейронів екстензорів. Ці впливи протилежні вестибулоспінальним. Завдяки руброспінальним впливам тонус м’язів мезенцефальної тварини більш рівномірніший, ніж у децереброваної (тонус флексорів та екстензорів більш збалансований). Тому, поза мезенцефальної тварини нормальна. Тобто, посилена антигравітаційна поза децереброваної тварини перетворюється в нормальну у мезенцефальної завдяки руброспінальним впливам. За участю рухових ядер СрМ здійснюються позо-тонічні рефлекси двох видів: 1.Випрямні – забезпечують відновлення порушеної пози. У тварин, що мають шию, випрямні рефлекси мають 2 фази: а) Рефлекс випрямлення голови – він починається з подразнення отолітових вестибулорецепторів у відповідь на порушення нормального положення голови в просторі (наприклад при падінні). Рівень замикання цього рефлексу – СрМ. Для його здійснення необхідна участь червоних ядер та РФ СрМ. По низхідних шляхах від цих структур інформація передається до мотонейронів спинного мозку Ù перерозподіл тонусу м’язів шиї Ù голова набуває нормальне положення в просторі (тім’ячком доверху). б) Рефлекс випрямлення тулуба – починається з подразнення пропріорецепторів шиї, що спричиняється рефлексом випрямлення голови, так як при цьому відновлюється нормальне положення голови у просторі, але порушується її нормальне положення відносно тулуба. Вхід інформації від пропріорецепторів шиї здійснюється на рівні спинного мозку Ù підйом до надсегментарних структур СрМ Ù обробка цієї інформації Ù передача до мотонейронів спинного мозку Ù до м’язів, які забезпечують відновлення нормального положення тулуба відносно голови у просторі. 2. Стато-кінетичні – забезпечують збереження пози рівноваги при русі з прискоренням. При русі з лінійним прискоренням збуджуються отолітові рецептори, а при русі з кутовим прискоренням – рецептори півколових канальців. Вхід цієї інформації в ЦНС здійснюється на рівні заднього мозку Ù підйом до структур СрМ Ù обробка та передача низхідними шляхами до мотонейронів спинного мозку Ù перерозподіл тонусу м’язів, який забезпечує збереження рівноваги під час руху з прискоренням. Важливим компонентом стато-кінетичних рефлексів є присінково-окорухові рефлекси Ù ністагм очей Ù більш продовжена фіксація предметів в полі зору під час руху Ù краща орієнтація в просторі Ù більш надійне збереження пози при русі. Ще раз підкреслимо, що рівень замикання випрямних та стато-кінетичних рефлексів – СрМ. Тому, їх можливо отримати у мезенцефальної тварини і неможливо у децереброваної чи спінальної. За рахунок обробки інформації від зорових та слухових рецепторів (за участю ядер чотирьох горбків) рухові ядра СрМ забезпечують здійснення орієнтаційних рефлексів – у відповідь на сильні звукові та світлові подразники мезенцефальна тварина повертає голову в бік подразника, насторожує вуха, в неї дещо посилюється тонус флексорів – тварина ніби готується до відповіді на цей подразник. Роль СрМ в здійсненні фазичних реакцій залежить від рівня організації тварини: - мезенцефальні земноводні (жаба) здатні до локомоцій – руху у просторі (стрибають, плавають); - кішки та собаки не здатні до локомоцій, але здійснюють локомоторні синергії – злагоджені рухи кінцівок, що нагадують акт ходьби: їх інтегрують структури СрМ разом з структурами заднього та спинного мозку. У людини СрМ не забезпечує формування нормальної антигравітаційної пози в спокої та підтримання її при русі. Для здійснення всіх цих функцій у людини необхідна участь кори великих півкуль (принцип кортикалізації функцій). 26. Мозочок, його функції, симптоми ураження . Мозочок (Мз) відіграє дуже важливу роль в координації рухів і виконує цю функцію на основі своїх аферентних та еферентних зв’язків. По аферентним волокнам сигнали поступають в кору мозочка, яка має складну будову та забезпечує тонку обробку поступившої інформації. Причому, кінцеву обробку інформації здійснюють грушоподібні клітини Пуркіньє – по відростках цих клітин (по їх аксонах) здійснюється еферентний вихід обробленої інформації з кори Мз. Далі, інформація здебільшого прямує до ядер Мз Ù по аксонах їх нейронів здійснюється вихід інформації за межі Мз. Аферентні зв’язки Мз: 1. Від рецепторів шкіри та пропріорецепторів Мз отримує інформацію по спіно-церебелярних шляхах; ця інформація забезпечує можливість контролю за рухом який здійснюється. 2. Від вестибулорецепторів (через вестибулярні ядра) – контроль за збереженням рівноваги при русі. 3. Від всіх рухових ядер стовбуру (ретикулярна формація, краєві ядра). 4. Від базальних ядер. 5. Від моторної і асоціативної кори (від моторної кори надходить копія команди до виконання руху, яка відправляється до мотонейронів; від асоціативної кори – інформація про ціль руху). 6. Від зорових та слухових рецепторів – контроль за рухами, які відбуваються. Еферентні зв’язки Мз: 1. З руховими ядрами стовбуру (ретикулярна формація, вестибулярні ядра, червоні ядра), через які Мз здійснює вплив на мотонейрони і на м’язи. 2. З базальними ядрами. 3. З асоціативною та моторною корою 4. З гіпоталамусом.
Таким чином, зв’язки Мз обширні і нерідко є двосторонніми (з корою, з базальними ядрами, з руховими ядрами стовбуру). Мз отримує інформацію про характер руху від рецепторів і при необхідності може швидко корегувати ці рухи та їх програму. При цьому різні відділи Мз здійснюють корекцію різних рухів.
Таким чином, кора древнього та старого Мз, отримуючи інформацію від пропріорецепторів, рецепторів шкіри про характер руху; від вестибулорецепторів – про збереження рівноваги, і при необхідності вносить корективи в грубі позо-тонічні рухи, чому передує передача інформації по шляхах МНС, які регулюють стан м’язів тулуба і проксимальних відділів кінцівок. Ця корекція дуже важлива для підтримання рівноваги (збереження пози) при русі.
а) в рух, який здійснюється (виходячи на мотонейрони спинного мозку через стовбурові рухові ядра); б) в рухову програму, передаючи інформацію по церебело-таламо-кортикальному шляху в рухову кору. Вихід до рухової кори і до стовбурових рухових ядер із кори проміжної частини Мз здійснюється через вставне ядро. За рахунок корекції рухів та програми Мз добивається їх повного співпадання (їх відповідності один одному).
Кора півкуль Мз пов’язана з базальними гангліями і з асоціативною корою. Завдяки цим зв’язкам вони формують програми рухів і Мз через зубчасте ядро передає їх в рухову кору для виконання.
Основні симптоми ураження Мз у людини. Мехпнізми їх виникнення. 1. Дистонія – порушення тонусу м’язів, що спричиняється порушенням впливу Мз на стовбурові рухові ядра (ретикулярна формація, вестибулярні ядра) Ù на мотонейрони спинного мозку Ù на м’язи тулуба та проксимальних відділів кінцівок. 2. Дизеквілібрія – перушення рівноваги. Пов’язана з випаданням коригуючих впливів Мз на вестибулярні ядра Дейтерса. 3. Атаксія – невпорядкованість рухів. Проявляється у вигляді: а) дисметрія – порушення амплітуди рухів; б) порушення направленості рухів. В основі атаксії лежать порушення координації м’язів агоністів та антагоністів. 4. Асинергія – порушення злагодженості рухів. В основі її лежить порушення координації скорочення м’язів-агоністів. 5. Астенія – швидка втомлюваність. Вона пов’язана з виконанням великої кількості “зайвих” рухів для досягнення мети. 6. Адіадохокінез – порушення синхронного виконання рухів, іх послідовності та швидкості. 7. Дисметрія – порушення рівномірності і амплітуди рухів. Дисартрія – порушення артикуляції мови. 27. Таламус, його функції. Таламус є структурою проміжного мозку і виконує, головним чином, сенсорні функції – за виключенням нюхового, в тамусі переключаються всі сенсорні шляхи, які йдуть в КГМ. В таламусі є біля 40 ядер, які розрізняються по своїм функціям. Серед них виділяють: 1. Сенсорні перемикаючі (специфічні) ядра – вони отримують інформацію від специфічних сенсорних шляхів, переробляють її і передають в сенсорні зони КГМ. Разом з цими зонами специфічні сенсорні перемикаючі ядра таламуса беруть участь в формуванні відчуттів. 2. Неспецифічні – вони отримують інформацію від ретикулярної формації стовбура мозку по шляхах больової чутливості. Вони передають інформацію до всіх зон КГМ, здійснюючи на неї неспецифічний активуючий вплив. 3. Асоціативні – отримують інформацію від специфічних сенсорних перемикаючих ядер і від неспецифічних ядер таламуса. Вони передають інформацію в асоціативні зони КГМ (фронтальна, тім’яна, вискова). Разом з даними ділянками кори, асоціативні ядра таламуса приймають участь в розпізнаванні образів. 4. Неспецифічні перемикаючі: а) лімбічні – отримують інформацію від гіпоталамуса та передають її в лімбічну кору. Вони беруть участь у формуванні емоцій та мотивацій. б) рухові – отримують інформацію від базальних гангліїв, мозочка, стовбурових рухових ядер і передають її в рухову кору. Беруть участь в забезпеченні рухових реакцій організму
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-26; просмотров: 890; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.113 (0.015 с.) |

 Включає кірково-спинномозковий шлях (tractus corticospinalis) та кірково-червоноядерно-спинномозковий шлях (tractus corticorubrospinalis). Вони проходять поруч в бокових канатиках спинного мозку, закінчуються переважно на вставних нейронах латеральних рухових ядер, які інервують м’язи дистальних відділів кінцівок. Обидва шляхи діють як синергісти та активують мотонейрони, що іннервують флексори та гальмують мотонейрони, що активують екстензори. Таким чином, за участю ЛНС регулюється переважно згинальні рухи дистальних відділів кінцівок, які у людей є найбільш тонкими та точними.
Включає кірково-спинномозковий шлях (tractus corticospinalis) та кірково-червоноядерно-спинномозковий шлях (tractus corticorubrospinalis). Вони проходять поруч в бокових канатиках спинного мозку, закінчуються переважно на вставних нейронах латеральних рухових ядер, які інервують м’язи дистальних відділів кінцівок. Обидва шляхи діють як синергісти та активують мотонейрони, що іннервують флексори та гальмують мотонейрони, що активують екстензори. Таким чином, за участю ЛНС регулюється переважно згинальні рухи дистальних відділів кінцівок, які у людей є найбільш тонкими та точними. Включає кірково-присінково-спинномозковий шлях (tractus corticovestibulospinalis) та кірково-сітчасто-спинномозковий шлях (tractus corticoreticulospinalis) – від моторної кори через медіальне ретикулярне ядро заднього мозку. Вони проходять поруч в передніх канатиках спинного мозку, закінчуються на мотонейронах медіальних рухових ядер, що інервують м’язи тулуба та проксимальних відділів кінцівок. Вплив цих шляхів антагогістичний – кірково-присінково-спинномозковий шлях активує мотонейрони, що інервують екстензори та гальмує мотонейрони, що інервують флексори, а кірково-сітчасто-спинномозковий шлях має протилежній вплив. Таким чином, за участю МНС відбуваєть регуляція грубих рухів тулуба та проксимальних відділів кінцівок.
Включає кірково-присінково-спинномозковий шлях (tractus corticovestibulospinalis) та кірково-сітчасто-спинномозковий шлях (tractus corticoreticulospinalis) – від моторної кори через медіальне ретикулярне ядро заднього мозку. Вони проходять поруч в передніх канатиках спинного мозку, закінчуються на мотонейронах медіальних рухових ядер, що інервують м’язи тулуба та проксимальних відділів кінцівок. Вплив цих шляхів антагогістичний – кірково-присінково-спинномозковий шлях активує мотонейрони, що інервують екстензори та гальмує мотонейрони, що інервують флексори, а кірково-сітчасто-спинномозковий шлях має протилежній вплив. Таким чином, за участю МНС відбуваєть регуляція грубих рухів тулуба та проксимальних відділів кінцівок.




 Після операції децеребрації у тварини виникає децеребраційна ригідність, яка у кішки проявляється у слідуючому: передні і задні кінцівки розігнуті; голова закинута назад за рахунок розгинання шиї; хвіст піднятий; тулуб розігнутий. Ця поза кішки пов’язана з гіпертонусом розгиначів. Позу децеребраційної тварини (децеребраційна ригідність) можна охарактеризувати як посилену позу антигравітації. Згадаємо, що спинний мозок забезпечував формування м’язового тонусу рефлекторної природи, але не забезпечував формування пози антигравітації із-за малої сили тонусу.
Після операції децеребрації у тварини виникає децеребраційна ригідність, яка у кішки проявляється у слідуючому: передні і задні кінцівки розігнуті; голова закинута назад за рахунок розгинання шиї; хвіст піднятий; тулуб розігнутий. Ця поза кішки пов’язана з гіпертонусом розгиначів. Позу децеребраційної тварини (децеребраційна ригідність) можна охарактеризувати як посилену позу антигравітації. Згадаємо, що спинний мозок забезпечував формування м’язового тонусу рефлекторної природи, але не забезпечував формування пози антигравітації із-за малої сили тонусу. В основі формування децеребраційної регідності лежать вестибулоспінальні впливи. Присінкові ядра Дейтерса тонічно активні, так як вестибулорецептори адаптуються дуже повільно. Від цих ядер по присінково-спинномозковим шляхам, що не перехрещуються, інформація поступає до мотонейронів, які інервують м’язи тулуба та проксимальних відділів кінцівок. При цьому a- та g-мотонейрони екстензорів активуються, а a- та g-мотонейрони флексорів, навпаки – гальмуються.
В основі формування децеребраційної регідності лежать вестибулоспінальні впливи. Присінкові ядра Дейтерса тонічно активні, так як вестибулорецептори адаптуються дуже повільно. Від цих ядер по присінково-спинномозковим шляхам, що не перехрещуються, інформація поступає до мотонейронів, які інервують м’язи тулуба та проксимальних відділів кінцівок. При цьому a- та g-мотонейрони екстензорів активуються, а a- та g-мотонейрони флексорів, навпаки – гальмуються.



 Древній та старий Мз (флокуло-нодулярна система та хробак) – отримують інформацію від вестибулорецепторів, пропріорецепторів та шкірних рецепторів. Кора цих відділів передає інформацію до ядра шатра і далі до вестибулярного ядра Ù до медіального ретикулярного ядра Ù до мотонейронів, що іннервують м’язи тулуба та кінцівок.
Древній та старий Мз (флокуло-нодулярна система та хробак) – отримують інформацію від вестибулорецепторів, пропріорецепторів та шкірних рецепторів. Кора цих відділів передає інформацію до ядра шатра і далі до вестибулярного ядра Ù до медіального ретикулярного ядра Ù до мотонейронів, що іннервують м’язи тулуба та кінцівок.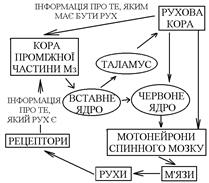 Проміжна частина кори Мз, як і кора древнього та старого Мз, отримує інформацію про те, який рух здійснюється. Від рухової кори сюди поступає інформація про те, який рух повинен відбуватися (копія рухової програми). Кора проміжної частини Мз працює як блок порівняння – порівнює інформацію про те, який рух повинен відбуватися, і про те, який рух проходить. При неспівпаданні цієї інформації Мз може вносити такі корективи:
Проміжна частина кори Мз, як і кора древнього та старого Мз, отримує інформацію про те, який рух здійснюється. Від рухової кори сюди поступає інформація про те, який рух повинен відбуватися (копія рухової програми). Кора проміжної частини Мз працює як блок порівняння – порівнює інформацію про те, який рух повинен відбуватися, і про те, який рух проходить. При неспівпаданні цієї інформації Мз може вносити такі корективи:


