Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Основные изменения нейрогуморальной регуляции при адаптации к повторным стрессорным воздействиямСодержание книги
Поиск на нашем сайте Многообразные экспериментальные исследования влияния повторных стрессорных воздействий позволяют выявить по меньшей мере три главных изменения, развивающиеся в организме при таких воздействиях: 1) адаптивное увеличение потенциальной мощности стресс-реализующих систем, 2) снижение степени включения таких систем, т. е. уменьшение стресс-реакции по мере повторения стрессорных ситуаций, и 3) снижение реактивности нервных центров и исполнительных органов к медиаторам и гормонам стресса — их своеобразная десенситизация. Увеличение мощности стресс-реализующих систем было наиболее убедительно продемонстрировано R. Kvetnansky и соавт. (1970) на примере системы биосинтеза катехоламинов в надпочечниках. Эти исследователи исходили из положения, что активность ключевого фермента биосинтеза катехоламинов тирозингидроксилазы возрастает при любом значительном увеличении расхода катехоламинов, и они предположили, что повторные стрессорные воздействия должны привести к увеличению активности тирозингидроксилазы, которая становится основой стационарного увеличения биосинтеза катехоламинов. Эксперименты показали, что уже после однократного иммобилизационного стрессорного воздействия, продолжавшегося 2,5 ч, активность тирозингидроксилазы в надпочечниках возрастает, и после 7 ежедневных сеансов иммобилизации она оказывается увеличенной более чем в 3 раза по сравнению с контролем. В дальнейшем повторные стрессорные воздействия, продолжавшиеся 6 нед, не оказали никакого дополнительного влияния на активность тирозингидроксилазы — установилось некоторое высокое плато активности фермента и биосинтеза катехоламинов, обеспечивающее стабильную гиперфункцию надпочечников при повторных стрессорных воздействиях. Прекращение стрессорных воздействий в соответственно прекращение указанной гиперфункции привело к резкому снижению активности фермента, которая в первые же 3 сут уменьшилась в 2 раза и на 14-е сутки вернулась к норме. Известно, что аналогичное увеличение активности тирозингидроксилазы в условиях компенсаторной гиперфункции надпочечников полностью подавляется ингибитором синтеза РНК актиномицином D или ингибитором синтеза белка циклогексимидом [Mueller R. et al., 1969]. Поэтому наблюдавшееся R. Kvetnansky и соавт. (1970) быстрое увеличение активности фермента при повторных стрессах следует рассматривать как результат генетически детерминированной активации биосинтеза этого фермента, а снижение его активности после прекращения стрессорных воздействий является результатом уменьшения скорости биосинтеза фермента до исходного уровня. Таким образом, динамика активности тирозингидроксилазы в надпочечниках при повторных стрессорных воздействиях почти полностью повторяет динамику массы органов, например сердца, при их гиперфункции и гипертрофии вследствие увеличенной нагрузки, а также динамику обратного развития гипертрофии после устранения нагрузки [Меерсон Ф. 3., 1975]. Такое совпадение неслучайно. Оно, очевидно, объясняется тем, что в основе обоих случаев «долговременной» адаптации лежит общий механизм — взаимосвязь между уровнем физиологической функции клеток и активностью их генетического аппарата. По существу этот фундаментальный механизм обеспечивает формирование структурной базы любой «долговременной» адаптации [Меерсон Ф. 3., 1963; 1984] и, в частности, адаптации к повторным стрессорным воздействиям. Весьма существенно, что увеличение активности тирозингидроксилазы в гипертрофированных надпочечниках при адаптации к повторным стрессорным воздействиям соответствует повышению содержания в них катехоламинов. Таким образом, физиологический резерв этого важного стресс-реализующего органа увеличен. При изучении влияния повторных стрессорных воздействий на метаболизм катехоламинов в мозгу было обнаружено аналогичное явление — потенциальная способность адренергических центров головного мозга осуществлять ресинтез и выделение катехоламинов в результате адаптации к стрессорным ситуациям возрастает [Geller Е. et al., 1965; Suthanthirorajan N., Subranmanyam S., 1983]. Таким образом, речь идет о генерализованном увеличении физиологической мощности важной стресс-реализующей системы организма. Торможение функции стресс-реализующих систем в условиях покоя выражается в замедлении обмена катехоламинов в мозге. Это проявляется в том, что у адаптированных животных снижение содержания катехоламинов при ингибировании их синтеза происходит значительно медленнее, чем у неадаптированных [Roth К. et al., 1982]. При эмоциональноболевом стрессе та же самая черта регуляции имеет более яркое выражение и заключается в том, что стресс-реакция на стрессорную ситуацию у адаптированных животных либо вообще не возникает, либо реализуется в незначительной степени. В табл. 8 представлены результаты исследований, проведенных совместно с В. В. Малышевым (1986), которые показывают это явление применительно к адренергическому (правая часть таблицы) и Таблица 8. Содержание катехоламинов и кортикостерона в плазме крови, сердце и надпочечниках крыс при длительном эмоционально-болевом стрессе (М±т)
гипофизарно-адреналовому (левая часть таблицы) звеньям стрессорной реакции. В этом исследовании крыс адаптировали к повторим эмоционально-болевым стрессорным воздействиям в двух режимах: 1) в режиме интенсивной адаптации, который предусматривал 35 эмоционально-болевых воздействий ежедневно, и 2) в щадящем режиме, при котором проводилось 10 эмоционально-болевых воздействий через день. Адаптированных животных подвергали затем однократному воздействию 6-часового эмоционально-болевого стресса (ЭБС). Рассмотрим два главных положения, вытекающих из материала таблицы. Первое из них состоит в том, что адаптация в обоих режимах привела к достоверному увеличению концентрации катехоламинов в надпочечниках и сердце и вместе с тем парадоксальным образом резко уменьшила степень мобилизации катехоламинов из этих органов при тяжелом стрессорном воздействии. Действительно, данные табл. 8 свидетельствуют, что у неадаптированных животных под влиянием такого стрессорного воздействия концентрация адреналина в надпочечниках уменьшалась почти в 3 раза, а концентрация норадреналина в сердце в 2 раза. У животных, предварительно адаптированных к стрессу, это уменьшение составляло всего окодо 20%, т. е. накопившийся в результате адаптации резерв катехоламинов при тяжелом стрессорном воздействии не мобилизовался. Второе положение заключается в том, что у адаптированных животных не реализовался гипофизарно-адреналовый эффект стресса. В контроле этот эффект состоял в том, что под влиянием 6-часового стрессорного воздействия концентрация кортикостерона возросла в надпочечниках в 2,5 раза, а в плазме крови в 3 раза. У адаптированных животных это явление полностью отсутствовало [Меерсон Ф. 3., Малышев В. В. и др., 1986]. Оба эти факта характеризуют угасание стрессорной реакции в результате адаптации к повторным стрессорным воздействиям. Они подчеркивают то, что адаптация ограничивает мобилизацию резерва стресс-реализующих систем, и вместе с тем совпадают с результатами, полученными первыми исследователями, изучавшими когда-то адаптацию организма к стрессорным воздействиям. Действительно, еще в 1976 г. К. Keim и Е. Sigg наблюдали, что через 30 мин после первого иммобилизационного стрессорного воздействия концентрация кортикостерона в крови возрастала в 10 раз, при повторном 30минутном воздействии подъем кортикостерона оказался еще выше, а при последующих воздействиях резко снижался; в итоге на 5-м иммобилизационном воздействии концентрация кортикостерона в крови возрастала всего лишь в 2 раза. Однако это уменьшение реакции гипофизарно-адреналовой системы на стрессорную ситуацию сочеталось с повышением ее фоновой активности — исходная концентрация кортикостерона в крови перед 5-м воздействием была увеличена более чем в 2 раза по сравнению с исходным фоновым уровнем. Эта фаза адаптации, характеризующаяся сочетанием уменьшенного ответа на стрессорную ситуацию с повышенной фоновой активностью гипофизарно-адреналовой системы, отмечена и в работах других исследователей [Виру А. А., 1974]. Можно предположить, что эта энергетически и структурно расточительная фаза «постоянной бдительности» является переходной на пути к более полной адаптации. Наиболее важные исследования по адаптации к неотвратимым стрессорным воздействиям были выполнены L. Mikulaj и соавт. [Mikulaj L. et al., 1974], которые показали, что при ежедневном введении формалина в кожу спины, а также при ежедневной иммобилизации у крыс первоначально возникает многократное повышение концентрации кортикостерона в крови. В дальнейшем эти подъемы постепенно уменьшаются и в ответ на 22—27-е стрессорное воздействие вообще не возникают. Снижение активации гипофизарно-адреналовой системы не зависит от истощения функциональных возможностей надпочечников, ибо введение адаптированным к стрессорной ситуации животным АКТГ вызывает у них большее повышение концентрации кортикостерона в крови, чем в контроле. Этот важный факт позволяет предположить, что в основе адаптации организма к многократному или длительному действию стрессорной ситуации лежит активация каких-то центральных тормозных механизмов, которые действуют на уровне мозга, и, несмотря на боль, тормозят возбуждение высших адренергических центров, выход рилизинг-факторов, АКТГ и таким образом предотвращают подъем содержания катехоламинов и кортикостерона в крови. Это представление соответствует известному из практики уменьшению циркуляторного компонента реакции — уменьшению подъема АД и тахикардии по мере привыкания к неотвратимым опасным ситуациям или болевым воздействиям. В интересной работе О. Desi-
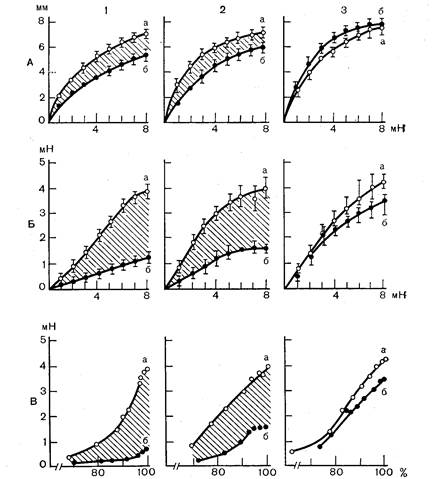
derate и соавт. (1974) эта же закономерность продемонстрирована по отношению к другому эффекторному органу — желудку. Показано, что у крыс при первом стрессорном воздействии — болевом раздражении лап без возможности избегания—возникла глубокая депресия секреции соляной кислоты в желудке, обусловленная, как указывалось ранее, сильным возбуждением адренергической регуляции при одновременном снижении тонуса блуждающего нерва. При действии той же самой стрессорной ситуации в четвертый раз такой депрессии не наблюдалось общая и свободная кислотность желудочного сока менялась мало. Существенно, что адаптивное угасание стресс-реакции при повторных стрессорных воздействиях проявляется не только ослаблением ее интенсивности — снижением гормональных ответов и реакции органов-мишеней, но также уменьшением длительности этих ответов. Выяснение этого положения связано с установленным еще Г. Селье фактом, что любой стресс сопровождается эозинопенией, вызванной избытком кортикостероидов [Selye H., 1960]. Оказалось, что при завершении стресс-реакции эозинопения не просто прекращается, но завершается пиком эозинофилов и, таким образом, время от завершения стрессорной ситуации до пика эозинофилов, т. е. длительности эозинопении, хороший критерий завершения стрессреакции. Чем короче этот период, тем ближе к завершению процесс адаптации к любому фактору и, в частности, к стрессорной ситуации [Малышев В. В. и др., 1982 а]. На рис. 8 показано, что после тяжелого 6-часового эмоционально-болевого стрессорного воздействия у неадаптированных животных пик эозинофилии наступает лишь через 42 ч; у животных, прошедших перед таким воздействием интенсивную адаптацию (35 раз по 40 мин ежедневно) тот же пик возникает уже через 36 ч, и, наконец, при использовании оптимальной адаптации (10 раз по 40 мин через день) эозинопения завершается и пик эозинофилов возникает уже через 18 ч после завершения тяжелого стрессорного воздействия. Таким образом, в результате адаптации к многократным коротким стрессорным воздействиям развивается состояние, которое, с одной стороны, выражается увеличением мощности стресс-реализующих систем, а с другой — постепенным уменьшением интенсивности и длительности стрессреакции даже на весьма интенсивные, заведомо повреждающие стрессорные ситуации. Для понимания этого явления надо иметь в виду, что в процессе адаптации, наряду с угасанием самой стресс-реакции, развивается своеобразное снижение чувствительности — десенситизация к стрессорным медиаторам и гормонам. Явление десенситизации при адаптации к повторным стрессорным воздействиям. Факт десенситизации продемонстрирован в исследованиях последних лет в форме снижения количества β-адренорецепторов и уменьшения чувствительности аденилциклазной системы мозга и периферических тканей к катехоламинам. В экспериментах некоторых исследователей это явление оказалось весьма значительным. Так, Т. Torda и соавт. (1981) оценивали состояние β-адренорецепторов в мозге по связыванию с меченым лигандом 3Н-дегидроаллренололом и обнаружили, что в результате повторных электроболевых воздействий количество рецепторов в гипоталамусе и коре больших полушарий уменьшилось на 50 и 25% соответственно; при этом их сродство к катехоламинам не изменялось. В более позднем и методически несомненно более совершенном исследовании Е. Stone и J. Platt (1982) это явление было подтверждено, и хотя количественно снижение плотности β-рецепторов в мозгу было меньшим, оно четко коррелировало с повышением резистентности крыс к стрессорным воздействиям. Рис. 9 отражает диннмику количества β-адренорецепторов в коре, гипоталамусе и стволе головного мозга в процессе адаптации, а именно при 14 повторных стрессорных воздействиях. Кривая на рис. 10 характеризует динамику анорексии, т. е. потери аппетита, и его восстановления в том же эксперименте. Видно, что оба процесса коррелируют между собой. Эта корреляция соответствует результатам ранних работ, которые свидетельствуют о том, что реакция на норадреналин в коре и гипоталамусе снижается только после повторных электроболевых раздражений, когда возрастает резистентность к анорексическому [Hitter S. et al., 1978; Stone E. A., 1979], поведенческому [Weiss J. et al., 1975] и летальному [Zigmond M., Harvey J., 1970] эффекту этих стрессорных воздействий. Представление о том, что десенситизация является реальным фактом адаптации к повторным стрессорным воздействиям, соответствует данным Т. Torda и соавт. (1981 а), которые показали, что повторный иммобилизационный стресс в течение недели уменьшает количество β-адренорецепторов не только в головном мозге, но и в сердце, а также α-адренорецепторов в селезенке. Этот факт совпадает с данными D. U'Prichard, R. Kvetnansky (1980), которые обнаружили уменьшение количества β-адренорецепторов в сердцах крыс после 14 иммобилизационных стрессов и систематического введения β-агониста изопротеренола. Снижение числа и эффективности функционирования адренорецепторов, т. е. адренореактивности, чем бы оно ни было вызвано, может стать важным фактором повышения резистентности исполнительных органов к стрессорному повреждению. Действительно, исследования, выполненные в нашей лаборатории в последнее время [Каткова Л. С. и др., 1984] показали, что резистентность миокарда животных различных генетических линий к стрессорному повреждению в высокой степени зависит от состо-
лния адренорецепторного аппарата сердечной мышцы и мембранного аппарата транспорта Са2+ в кардиомиоцитах. В этой работе сопоставляли резистентность к длительному иммобилизационному стрессу сердца и желудка крыс трех генетических линий, а именво нормотензивных линий Wistar—Kyoto (WKY) и Август и крыс со спонтанной наследственной гипертонией (SHR). Для миокарда крыс последней пинии доказано уменьшение хронотропного и инотропного эффекта катехоламинов и их синтетических аналогов, действующих через |3-адренорецепторы [Hausell В. В., Alpert N. Р., 1976]. Оно обусловлено уменьшением количества [Limas С., Limas С., 1978] или сродства [Giachetti A. et al., 1979] к катехоламинам самих (5-адренорецепторов и снижением содержания циклического аденозинмонофосфата (цАМФ) в сердечной мышце [Dhalla N. et al., 1978]. Вполне возможно, что аналогичная ситуация имеется не только в сердце, но и в других органах, в частности в желудке. Все животные подверглись длительному 6-часовому пммобилизационному стрессу, реакция на который оказалась различной у крыс разных генетических линий. У крыс линии WKY средняя длина язвенных поражений слизистой в расчете на один желудок составляла 18 мм и одновременно наблюдались выраженные нарушения сократительной функции миокарда. У крыс линии Август язвенные поражения слизистой оболочки желудка отсутствовали, но нарушения сократительной функции миокарда были выражены примерно также, как и WKY. Наконец, у крыс линии SHR полностью отсутствовали поражения слизистой желудка и нарушения сократительной функции миокарда. Детальное изучение сократительной функции изолированного предсердия позволило сопоставить растяжимость миокарда, развиваемое напряжение и эффективность реализации механизма Старлинга у крыс изучаемых линий и влияние на эти параметры иммобилизационного стресса. Из рис. 11 видно, что до стрессорного воздействия основные параметры, характеризующие сократительную функцию сердечной мышцы, а именно: растяжимость миокарда, сила сокращения, развиваемая на плато кривой Старлинга, показатель эффективности реализации механизма Старлинга, у животных всех трех генетических линий оказались одинаковыми. Влияние перенесенного стресса на те же параметры, напротив, оказалось существенно различным у животных разных генетических линий. Действительно, у крыс линии WKY в соответствии с ранее полученными для крыс линии Вистар данными [Воронцова Е. Я., Пшенникова М. Г., Меерсон Ф. 3., 1982] после стресса наблюдалось существенное снижение растяжимости, наиболее определенно выраженное при небольшой внешней нагрузке, и одновременно произошло снижение развиваемого напряжения и величины показателя эффективности реализации механизма Старлинга более чем в два раза (р<0,05). Аналогичная депрессия растяжимости и показателей сократительной функции развивалась под влиянием стресса у животных линии Август. У гипертензивных животных линии SHR стресс, напротив, не вызывал каких-либо достоверных изменений растяжимости, эффективности реализации механизма Старлинга и развиваемого напряжения. Таким образом, по показателям растяжимости и сократительной функции у животных линии SHR резистентность миокарда к повреждающему действию иммобилизационного стресса оказа-
лась существенно повышенной по сравнению с животными других линий. В связи с этим представляют интерес данные исследований последних лет о возможном механизме десенситизации. По этому поводу имеется две группы работ, которые связывают явление десенситизации с изменением липидного бислоя клеточных мембран нейронов и исполнительных органов, т. е. по существу с изменениями липидного микроокружения адренорецепторов при повторных стрессорных воздействиях. Так, Т. Torda и соавт. (1981 а) пришли к заключению, что во время десенситизации в мембранах происходит разрушение фосфатидилхолина и накопление продуктов его распада — лизофосфатидилхолина и арахидоновой кислоты. Ингибитор фосфолипазы А блокирует этот процесс и делает невозможной десенситизацию in vivo и in vitro в мозге и сердце. Другая работа по тому же вопросу опирается на ранее установленный нами факт, что острый эмоционально-болевой стресс вызывает активацию ПОЛ в мозге и исполнительных органах [Меерсон Ф. 3., 1981]. Развивая это положение, Л. Л. Прилипко с соавт. (1983) показали, что после острого эмоционально-болевого стресса связывание меченого лиганда 3Н-алпренолола оказывается уменьшенным на 60%. Это явление не сопровождается изменениями фосфолипидного или жирнокислотного состава биомембран, но при этом наблюдается увеличение содержания продуктов ПОЛ — гидроперекисей липидов и шиффовых оснований в 2—3 раза. Была высказана мысль, что именно активация ПОЛ в биомембранах может играть роль в явлениях десенситизации. Таким образом, оба предположения связывают адаптивную десенситизацию с изменением липидного обмена биомембран и, по-видимому, требуют критического анализа и развития, так как упомянутые изменения должны быть более выражены при остром стрессе, чем при привыкании к нему, между тем десенситизация и снижение адренореактивности, по-видимому, адаптивный сдвиг. В 1981 г. появились радиоиммунологические данные, свидетельствующие о том, что явление десенситизации не ограничивается уменьшением числа адренорецепторов, а распространяется также на цитоплазматические рецепторы кортикостероидов, число которых в кардиомиоцитах крыс закономерно уменьшалось после 10 кратковременных стрессорных воздействий (плаванье с грузом) [Eller A. et al ., 1981]. Это говорит о том, что адаптация к стрессорным ситуациям среды обеспечивается тем, что реакция стресс-реализующих систем, несмотря на увеличившуюся их мощность, почему-то не реализуется — стресс-реакция в процессе адаптации угасает. Одновременно чувствительность органов-мишеней (от постсинаптических нейронов до исполнительных органов) снижается — возникает десенситизация.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 473; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.62 (0.016 с.) |






 Рис. 9. Число ß-адренорецепторов в головном мозге крыс в процессе адаптации к стрессу. А — гипоталамус; Б — кора головногомозга; В — ствол мозга, 1 — контроль, принятый за 100%; 2 — адаптация. Ордината — число мест связывания 3НДНА, % от контроля; абсцисса — день повторных стрессорных воздействий. Для каждой точки n=6 [по Е. Stone, J. Platt, 1982].
Рис. 9. Число ß-адренорецепторов в головном мозге крыс в процессе адаптации к стрессу. А — гипоталамус; Б — кора головногомозга; В — ствол мозга, 1 — контроль, принятый за 100%; 2 — адаптация. Ордината — число мест связывания 3НДНА, % от контроля; абсцисса — день повторных стрессорных воздействий. Для каждой точки n=6 [по Е. Stone, J. Platt, 1982].