
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Методы исследования эволюцииСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте Экологическая характеристика популяций Популяции формируются исторически в известных экологических условиях, поэтому каждая популяция характеризуется экологической определенностью - распространена на своем популяционном ареале в пределах ареала вида, имеет большую или меньшую численность особей, половую и возрастную структуру, свою популяционную динамику.
Эволюционно-генетическая характеристика популяции Генетически популяция характеризуется ее генофондом (аллелофондом), который представлен совокупностью аллелей, образующих генотипы организмов данной популяции. Генофонды природных популяций отличает наследственное разнообразие (генетическая гетерогенность или полиморфизм), генетическое единство, динамическое равновесие доли особей с разными генотипами. Наследственное разнообразие связано с наличием в генофонде одновременно различных аллелей отдельных генов. Первично оно создается мутационным процессом. Мутации, будучи обычно рецессивными и не влияя на фенотипы гетерозиготных организмов, сохраняются в генофондах популяций в скрытом от естественного отбора состоянии. Накапливаясь, они образуют "резерв наследственной изменчивости". Благодаря комбинативной изменчивости этот резерв используется для создания в каждом поколении новых комбинаций аллелей. Объем такого резерва огромен. Так, при скрещивании организмов, различающихся по 1000 локусам, каждый из которых представлен десятью аллелями, количество вариантов генотипов достигает 10100, что превосходит число электронов во Вселенной. Число локусов (генов) у человека превышает эту цифру 10100 на 2-3 порядка. Генетическое единство популяции обусловливается достаточным уровнем панмиксии. В условиях случайного подбора скрещивающихся особей источником аллелей для генотипов организмов последовательных поколений является генофонд популяции. В пределах генофонда популяции доля генотипов, содержащих разные аллели одного гена, при соблюдении некоторых условий из поколения в поколение не изменяется. Эти условия описываются основным законом популяционной генетики, сформулированным в 1908 г. английским математиком Дж. Харди и немецким врачом-генетиком Г. Вайнбергом [показать]. Популяция, генофонд которой не изменяется в ряду поколений, называется менделевской. Генетическая стабильность менделевских популяций ставит их вне процесса эволюции, так как в таких условиях приостанавливается действие естественного отбора. Выделение менделевских популяций имеет чисто теоретическое значение. В природе эти популяции не встречаются, т.к. генетическая инертность преодолевается благодаря мутациям, притоку в популяцию или оттоку из нее особей с определенными генотипами и отбору. Т.о., отклонение от закона Харди-Вайнберга, описывающего условия генетической стабильности популяции, стало подтверждением эволюционного процесса [показать]. Формы искусственного отбора. Тщательное изучение явления искусственного отбора позволило Дарвину выделить две формы этого отбора: бессознательный и методический. Бессознательный отбор — это отбор, при котором не ставится цель создания нового сорта или породы. Люди сохраняют лучших, на их взгляд, особей и уничтожают (выбраковывают) худших. В сельских районах и в наше время хозяева осуществляют бессознательный отбор с курами, собаками, голубями. Бессознательный отбор возник давно, с момента приручения собаки. Бессознательный отбор ведет к изменению растений и животных, к улучшению пород и сортов, к созданию новых местных пород и сортов. Нужный результат при этом отборе формируется медленно, но он может быть впечатляющим. Так, во время археологических раскопок в Перу были обнаружены зерна кукурузы в 34 раза большего размера, чем современные. Предки таджиков (согды) выращивали абрикос, плоды которого содержали до 70 % сахара. Высыхая на деревьях, эти плоды не опадали. Методический отбор — это отбор, осуществляемый человеком по определенному плану, с определенной целью — создания породы или сорта. Для методического ИО характерны следующие особенности: · ставится цель: селекционер решает, какие признаки надо изменить и в какую сторону, т. е. определяется направление отбора (яйценоскость, мясистость, красивый гребень, красивый хвост, красивое оперение); · составляется план создания породы (сорта): какие породы (сорта) ив каком порядке нужно скрещивать, какие виды скрещивания нужно применять; · создаются специальные условия жизни; · методический отбор - это творчество. АЛЛОПАТРИЧЕСКОЕ ВИДООБРАЗОВАНИЕ Аллопатрическое (географическое) видообразование, как правило, происходит медленно и даёт виды, отличающиеся по морфофизиологическому критерию от вида-родоначальника.
1. 2. 3. Механизм аллопатрического видообразования: 4. 5. 6. 7. 8. Возникновение географической преграды (реки) приводит к возникновению изолятов — географически изолированных популяций. 9. 1. Каждый изолят будет эволюционировать независимо от других популяций. В изолированных частях будут накапливаться новые генотипы и фенотипы. Особи в разных частях ранее единого ареала могут изменить свою экологическую нишу. 2. 3. 4. 5. При таких исторических процессах степень расхождения групп может достигнуть видового уровня.
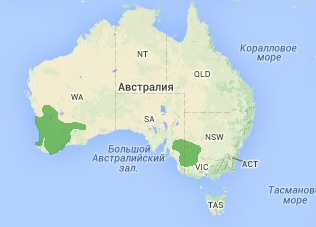
В Австралии обитает роскошный горный попугай Polytelis anthopeplus (Lear). В условиях засушливого периода единый ареал разделился пустыней Викторией на два разобщённых между собой ареала — на юго-востоке и юго-западе Австралии. В настоящее время оба подвида отличаются по своему поведению и местообитанию (этологический и экологический критерии). Восточно-австралийская популяция (Polytelis anthopeplus anthopeplus) избегает соседства с человеком и держится в степях с отдельно стоящими колками густых кустарников. Западно-австралийская популяция горного попугая (Polytelis anthopeplus monarchoides), наоборот, охотно заселяет культурный ландшафт и настолько увеличилась в числе, что наносит ощутимый вред полям пшеницы.
Так как между географическими популяциями попугаев не происходит обмена генетической информацией, в будущем возможно возникновение репродуктивной изоляции и, следовательно, видообразования. Большинство видов, особенно животных, возникают именно аллопатрическим путем. СИМПАТРИЧЕСКОЕ ВИДООБРАЗОВАНИЕ Симпатрическое (экологическое) видообразование происходит на одной территории и связано с появлением в популяции нескольких групп, особи которых не могут скрещиваться между собой. Симпатрический путь видообразования у паразитов часто связан с освоением популяцией новых хозяев. Анализ генного состава и межхромосомных различий между человеком и человекообразными обезьянами даёт повод предположить, что разделение этих двух ветвей могло идти симпатрическим путем. В природе разные виды африканских цихлид обычно не скрещиваются между собой. Репродуктивная изоляция обеспечивается в основном брачными предпочтениями. По-видимому, одним из главных факторов, обеспечивших быстрое видообразование у цихлид, был половой отбор.
Также симпатрическим путём возник новый вид цихлиды в озере Апойо, Никарагуа — учёным удалось показать, что и исходный вид, и новый являются потомками одной когда-то попавшей в это озеро самки (или немногих самок, но митохондриальные ДНК их не сохранились). Репродуктивная изоляция двух видов ворон — серой и чёрной — обеспечивается мутациями участка хромосомы, отвечающего за цвет оперения и поведение птицы. На границе ареалов происходит гибридизация видов.
Серая и чёрная вороны
У видов с половым размножением симпатричеcкое видообразование без географической изоляции происходит довольно редко, т. к. новые мутации либо распространяются в популяции (при возможности скрещивания с особью-мутантом), либо уходят вместе с гибелью носителя (при невозможности скрещивания с особью-мутантом). Однако у организмов, размножающихся бесполым путем, в том числе у высших растений с вегетативным размножением, один мутант (например, полиплоид), достаточно отличающийся от родительской популяции, чтобы быть генетически изолированным, может дать начало новому виду. Вариантами симпатрического видообразования являются полиплоидия и гибридизация. Полиплоидия — тип геномной мутации: увеличение числа наборов хромосом в клетках организма, кратное гаплоидному (одинарному) числу хромосом. Так, разные виды пшениц составляют ряд с наборами 14, 28, 42 хромосомы. В клетках дикого хлопчатника 26 хромосом, культурного — 52. Гибридизация — скрещивание организмов разных видов, т. е. объединение различных геномов в одной особи (гибриде). Культурная слива возникла путем гибридизации тёрна с алычой. Другим примером гибридогенного вида является рябинокизильник, распространённый в лесах центральной Сибири.
Рябинокизильник — природный гибрид рябины и кизила. ИЗОЛИРУЮЩИЕ МЕХАНИЗМЫ
Факторы, мешающие скрещиванию особей (изолирующие механизмы): · экологическая изоляция — особи различных групп предпочитают различные условия местообитания; · сезонная изоляция — оплодотворение у особей разных групп происходит в различное время; · этологическая (поведенческая) изоляция — особи разных групп имеют различные ритуалы ухаживания; · морфологическая (механическая) изоляция — репродуктивные органы особей разных групп делают невозможным перекрёстное оплодотворение. КОЛЬЦЕВЫЕ КЛИНЫ
гибридогенное видообразование В основе постзиготической изоляции, то есть сниженной жизнеспособности или плодовитости гибридов, лежит несовместимость геномов двух разных видов. В каких-то случаях они просто не могут работать вместе, и формирование организма нарушается. Однако, если у такого гибрида происходит геномная мутация, вызывающая удвоение числа хромосом, то мейоз протекает нормально и даёт нормальные половые клетки. При этом гибридная форма приобретает способность к размножению и утрачивает возможность скрещивания с родительскими формами. Таким образом, полиплоидизация открывает путь к гибридизации видов. Это часто происходит у растений. Например, известны виды пшеницы с 14, 28 и 42 хромосомами, виды роз с 14, 28, 42 и 56 хромосомами и виды фиалок с числом хромосом, кратным 6 в интервале от 12 до 54. Гексаплоидный (6n) геном мягких пшениц образовался путём объединения диплоидных геномов двух предковых видов пшениц и одного вида близкого рода Эгилопс (Aegilops). По некоторым данным, гибридогенное происхождение имеют не менее трети всех видов цветковых растений. Кроме того, межвидовые гибриды растений могут размножаться вегетативным путем и распространяться за счет этого.
Как правило, репродуктивная изоляция сначала возникает на постзиготических уровнях, а затем, поскольку тратить время и ресурсы на ухаживание, спаривание, гнездование при слабой жизнеспособности и плодовитости потомства невыгодно, возникают презиготические барьеры — например, поведенческие. Однако в случае географического видообразования изоляционные барьеры могут успеть сформироваться не в полной мере — скрещивание всё равно не происходит из-за наличия территориального разобщения. Поэтому от многих видов, изолированных географически, можно в неволе или искусственно получить потомство. Например, при скрещивании львов и тигров (разные виды одного рода Panthera) рождаются (в зависимости от пола родителей) лигры и тигрольвы, или тайгоны — здоровые и даже частично плодовитые животные. Темпы видообразования
Подобно формам видообразования столь же разнообразны и его темпы, варьирующие от относительно медленного и постепенного преобразования генофондов географически обособленных популяций при «классической» аллопатрической модели до чрезвычайно быстрого (в процессе смены всего нескольких поколений) обособления генофонда популяции основателей при «генетической революции» и действии катастрофического отбора, или столь же быстрого формирования нового вида при полиплоидизации и стасигенезе. В 1972 г. Н.Элдридж и С.Гулд выдвинули получившую широкую известность гипотезу так называемого «прерывистого равновесия», или «пунктуализма», которую они считают альтернативной классическим представлениям дарвинизма о постепенном («градуалистском») видообразовании. Согласно этой гипотезе история любой филетической линии состоит из чередования длительных периодов эволюционной стабильности — до 99 \% времени существования вида — и относительно кратких периодов видообразования: каждый акт видообразования охватывает около 10000—100000 лет. В качестве основного аргумента в пользу своей гипотезы Гулд и Элдридж использовали широко распространенное явление прерывистости филетических линий в палеонтологической летописи, обычно объясняемое фрагментарностью палеонтологических данных и приводящее к описанию дискретных во времени палеонтологических видов. Сторонники пунктуализма видят основные механизмы ускоренного видообразования в процессах генетических революций, происходящих в периферических изолятах; в сериях макромутаций (особенно в мутациях генов-регуляторов и хромосомных перестройках) и в так называемом «видовом отборе». Этот термин используется для обозначения отбора по признакам, инвариантным (единообразным) для вида в целом. В этом случае виды рассматривают как аналоги особей, причем сторонники пунктуализма полагают, что «видовой отбор» должен приводить к быстрому замещению хуже приспособленных по данному признаку видов лучше приспособленными. Однако необходимо помнить, что отбор — это вероятностный процесс, эффективный лишь при достаточно больших выборках. Число конкурирующих видов обычно значительно меньше минимального числа особей в популяции, начиная с которого отбор становится более эффективным, чем дрейф генов. Отбор по инвариантным видовым признакам, конечно, может происходить, но он базируется не на видах как целостных единицах, а на большом числе составляющих их особей. Поэтому такой отбор ничем принципиально не отличается от обычного естественного отбора и не требует выделения в особую категорию. Проверка многочисленных палеонтологических данных о характере видообразования в различных филетических линиях, проделанная целым рядом ученых, показала, что в палеонтологических рядах форм можно обнаружить и «градуалистское», и «пунктуалистское» видообразование. Вообще же, «разрешающая способность» палеонтологической летописи, как правило, совершенно недостаточна для того, чтобы по палеонтологическим данным можно было с достаточной достоверностью судить о характере процессов видообразования, поскольку «разрывы» между предками и потомками могут быть связаны как с высокими темпами видообразования, так и с неполнотой палеонтологической летописи. Ускоренное видообразование, утверждаемое сторонниками пунктуализма, вообще говоря, вполне возможно, и механизмы его известны (это «генетические революции», стасигенез, поли-плоидизация, гибридогенез). Однако неверно главное положение гипотезы прерывистого равновесия, что это — единственная (или резко преобладающая) форма эволюции, чему противоречат многочисленные данные по современным и ископаемым формам. Хотя сама концепция пунктуализма в целом не выдержала критики, дискуссии с ее сторонниками в 70—80-е гг. способствовали развитию представлений о многообразии форм и темпов видообразования.
Направления эволюции Направление эволюции каждой систематической группы определяется взаимоотношениями между особенностями среды, в которой протекает эволюция данного таксона, и его генетической организацией, которая сложилась в ходе его предшествующей эволюции.
Ярким примером дивергенции может служить изменение конечностей млекопитающих в ходе их приспособления к разным условиям среды. Рука человека, крыло летучей мыши, копыто лошади, лапа медведя, ласта морского льва, плавник кита – это все результаты дивергенции. Они возникли путем длительного естественного отбора мелких случайных уклонений в строении передней конечности общего предка млекопитающих. Его потомки заняли со временем разные экологические ниши. Этот процесс направлялся естественным отбором. В ходе эволюции накапливались изменения в генах, контролирующих детали развития конечностей, но начальные стадии их формирования в онтогенезе каждого из столь разных видов очень сходны и регулируются сходными генами. С труктуры и органы, имеющие общее происхождение называют гомологичными. Гомология конечностей у позвоночных столь выражена, что сходные элементы прослеживаются спустя сотни миллионов лет после начала дивергенции. С развитием молекулярной генетики и расшифровкой отдельных генов и целых геномов стало ясно, что дивергенция – это основное направление эволюции. Было показано, например, что различия в формировании конечностей у разных таксонов позвоночных (рыб, птиц, млекопитающих) обусловлены дивергенцией основных генов, направляющих этот процесс. Появилась возможность выяснить, какой из факторов эволюции обусловливал дивергенцию тех или иных элементов генов на разных этапах эволюции. Гены, контролирующие давно сложившиеся функции, например, гены домашнего хозяйства, дивергировали очень медленно, находясь под постоянным давлением стабилизирующего отбора. При этом дивергенция, как правило, ограничивалась нейтральными, синонимическими заменами нуклеотидов и, следовательно, была обусловлена дрейфом генов. В то же время дивергенция эволюционно молодых генов имела явно приспособительный характер. Например, эволюция генов опсинов – фоторецепторов сетчатки глаза - была сопряжена с большим количеством несинонимических замен нуклеотидов. Это привело к расширению спектра воспринимаемых цветов. В данном случае дивергенция на уровне генов направлялась естественным отбором.
Конвергенция (схождение признаков) наблюдается в тех случаях, когда неродственные таксоны приспосабливаются к одинаковым условиям. О конвергенции говорят в тех случаях, когда обнаруживается внешнее сходство в строении и функционировании какого-либо органа, имеющего у сравниваемых групп живых организмов совершенно разное происхождение. Например, крыло стрекозы и летучей мыши имеют общие черты в строении и функционировании, но формируются в ходе эмбрионального развития из совершенно разных клеточных элементов и контролируются разными группами генов. Такие органы называют аналогичным. Они внешне сходны, но различны по происхождению, они не имеют филогенетической общности. Сходство в строении глаз у млекопитающих и головоногих моллюсков — другой пример конвергенции. Они возникли независимо в ходе эволюции и формируются в онтогенезе из разных зачатков. Конвергенция затрагивает не только отдельные органы. Неродственные виды животных, эволюционируя в сходных условиях среды, занимая сходные экологические ниши, часто приобретают совершенно поразительное сходство. В Австралии, Африке и Южной Америке обитает множество муравьев и термитов. Ниша муравьедов на разных континентах была заполнена разными неродственными видами млекопитающих. В Австралии и Океании - это сумчатый муравьед и ехидна, которая принадлежит подклассу яйцекладущих млекопитающих. В Африке муравьев и термитов сделали своей основной пищей трубкозубы, дальние родственники слонов, и панголины, принадлежащие к отряду ящеров. В Южной Америке эту нишу заняли броненосцы и муравьеды – представители отряда неполнозубых (рис. XI.21.1.). Последний общий предок этих животных существовал более 100 миллионов лет назад. Ближайшие родственники каждого из них абсолютно на них непохожи. Только выбор экологической ниши и отбор на совершенствование приспособлений к этой нише сделал их внешне похожими друг на друга. Все они имеют длинный липкий язык и вытянутый хоботком нос, что позволяет им извлекать муравьев и термитов из их укрытий. Но при этом фундаментальные свойства анатомии этих видов позволяют безошибочно относить их к разным таксонам, несмотря на их конвергентное сходство. Данные молекулярно- генетического анализа убедительно показывают, что эти виды разделяют десятки миллионов лет независимой эволюции.
Параллелизм - такое эволюционное явление, когда сходство организмов, относящихся к разным таксонам, основано на сходных изменениях одних и тех же гомологичных структур. В качестве примера морфологического параллелизма можно указать на большое сходство формы тела у акулы, ихтиозавра (вымершей рептилии) и дельфина. Предки ихтиозавров и предки дельфинов были наземными животными. По мере освоения ими водной среды эволюция осевого скелета этих животных шла на основе гомологичных зачатков позвоночного столба в сходных условиях. Сходные направления естественного отбора привели к возникновению форм животных с близкими гидродинамическими характеристиками. Примером функционального параллелизма может служить возникновение крыльев птеродактилей, птиц и летучих мышей. Несмотря на то, что они устроены по-разному, они, очевидно, являются модификациями одного и того же органа - передней конечности, возникли в результате естественного отбора в сходных экологических нишах и выполняют одинаковые функции. Другой причиной параллелизма может быть относительно высокая вероятность сходных мутаций одних и те же генов. Согласно закону гомологических рядов Н.И.Вавилова родственные виды характеризуются сходными рядами наследственной изменчивости. Так, например, для многих видов бабочек характерно наличие на крыльях глазков разной формы, величины и окраски. При этом ближайшие родственники этих бабочек этих глазков не имеют. Очевидно, что этот признак возникал независимо у разных видов, а не был унаследован от общих предков. От них все виды бабочек унаследовали гены, мутации которых приводят к образованию глазков. Примером такого же рода является неоднократно и явно независимое появление саблезубых форм. Наиболее известен саблезубый тигр. Неудивительно, что были и другие саблезубые кошки. Но в палеонтологической летописи Австралии мы также находим саблезубого сумчатого волка. Общие и частные приспособления. Вопросы о возможных путях эволюционного процесса разработал А. Н. Северцов. Один из главных таких путей, по Северцову, — ароморфоз (арогенез), или возникновение в ходе эволюции приспособлений, которые существенно повышают уровень организации живых организмов и открывают перед ними совершенно новые эволюционные возможности. Такими приспособлениями были, например, возникновение фотосинтеза, полового размножения, многоклеточности, легочного дыхания у предков амфибий, амниотических оболочек у предков рептилий, теплокровности у предков птиц и млепитающих и др. Ароморфозы — естественный результат эволюционных процессов. Они открывают возможности для освоения видами новых, прежде недоступных сред обитания. Ароморфозы не возникают мгновенно, при появлении они практически неотличимы от обычных адаптаций. Лишь по мере их эволюционной «шлифовки» естественным отбором, согласования с многочисленными признаками организма и широкого распространения у многих видов они становятся ароморфозами. Например, появление легочного дыхания у древних обитателей пресных водоемов не изменило кардинально образа их жизни, уровня организации и т. д. Однако в результате возникновения этой адаптации появилась возможность для освоения суши — обширной среды обитания. Эта возможность была активно использована в последующей эволюции, появились многие тысячи видов амфибий, рептилий, птиц и млекопитающих, заполнивших разнообразные ниши обитания. Поэтому обретение позвоночными легких — крупный ароморфоз, приведший к повышению уровня организации многих видов. Возникают и менее крупные ароморфозы. В эволюции млекопитающих их было несколько: появление шерстного покрова, живорождение, вскармливание детенышей молоком, приобретение постоянной температуры тела, прогрессивное развитие мозга и др. Высокий уровень организации млекопитающих, достигнутый благодаря перечисленным ароморфозам, позволил им освоить новые среды обитания. Кроме такого крупного преобразования, как ароморфоз, в ходе эволюции отдельных групп возникает большое количество мелких приспособлений к определенным условиям среды. Такие приспособления А. Н. Северцов назвал идиоадаптациями.
Идиоадаптации — это приспособления организмов к окружающей среде без принципиальной перестройки биологической организации. Пример идиоадаптации — разнообразие форм у насекомоядных млекопитающих, разные виды которых, имея общий исходный уровень организации, смогли приобрести свойства, позволившие им занять разные места обитания в природе. Среди многих паразитических организмов мы обнаруживаем явление общей дегенерации, то есть резкого упрощения организации, связанного с исчезновением целых систем органов и функций. У крабов известен паразит саккулина, имеющий вид мешка, набитого половыми продуктами, и обладающий пищеварительной системой, пронизывающей тело хозяина. Его далекие предки принадлежали к усоногим ракам и обитали на дне моря, прикрепляясь к подводным камням. Их потомки постепенно перешли к паразитическому способу существования, утратив во взрослом состоянии почти все органы. Несмотря на то, что общая дегенерация приводит к значительному упрощению организации, виды, идущие по этому пути, могут увеличивать численность и ареал. Соотношение путей и темпы эволюции. Пути эволюции органического мира либо сочетаются друг с другом, либо сменяют друг друга, причем ароморфозы происходят значительно реже идиоадаптации. Но именно ароморфозы определяют новые этапы в развитии органического мира. Возникнув путем ароморфоза, новые, высшие по организации группы организмов занимают другую среду обитания. Далее эволюция идет по пути идиоадаптации, иногда и дегенерации, которые обеспечивают организмам освоение новой для них среды обитания. Законы эволюции на уровнях выше видового вытекают из принципов, работающих на популяционно-видовом уровне. Однако, чем дальше в прошлое, тем менее определенно мы видим детали эволюционного процесса, тем менее точно мы можем датировать ископаемые останки. Когда их возраст около 1 млн. лет, точность датировки колеблется в пределах нескольких тысяч лет. Ископаемые останки динозавров, которые жили около 120 млн. лет назад, ученые могут датировать с точностью до десятков или сотен тысяч лет. Для эпохи трилобитов (400 млн. лет назад) максимальная точность датировки падает до полумиллиона лет. Чем дальше в глубь времен, тем меньше мы находим ископаемых останков. Изменения морфологии, которые в далеком прошлом, происходили в течение десятков и сотен тысяч лет, десятков и сотен тысяч поколений, иногда кажутся в палеонтологической летописи, практически мгновенными. Это создает большие трудности в изучении конкретных механизмов эволюционных изменений имевших место в далеком. Иногда создается иллюзия, что тот или иной таксон образовался в результате резкого изменения организации предковой формы. Некоторые ученые пытались объяснить такие изменения появлением единичных крупных мутаций. Это предположение не выдерживает критики. Любой фенотипический признак есть результат согласованного действия множества генов. Чем сильнее фенотипический эффект мутации любого гена, вовлеченного в эти взаимодействия, тем выше вероятность, что она окажется гибельной для ее носителей. Новый крупный таксон, например, класс птиц, не мог возникнуть сразу, этому предшествовал длительный процесс, причем существовала серия промежуточных видов, у которых темпы эволюции отдельных органов и структур резко отличались. Развитие одних органов шло с большим опережением, другие развивались медленно. На примере ископаемого существа — археоптерикса видно, что по многим признакам (наличие зубов, несросшиеся позвонки хвоста, строение грудины, и др.) этот вид очень похож на рептилий. В то же время у археоптерикса есть и признаки, типичные только для птиц: перья, вилочковая кость, большие глаза, крылья и т. д. Обнаруживается также немало промежуточных признаков. Таким образом, археоптерикс обладает значительной мозаичностью организации — новые, типичные для птиц свойства сочетаются с большим количеством черт организации рептилий. Если бы развитие птиц в ходе эволюции не состоялось, то, обнаружив останки археоптерикса, ученые без колебаний отнесли бы этот вид к рептилиям. Освоение новой среды обитания сопровождается интенсивным естественным отбором по некоторым признакам. Для предков птиц таким признаком было крыло и структуры с ним прямо связанные. Соответствующий орган, структура или функция эволюционируют очень быстро, тогда как другие отстают. Таким образом, отмечается не взаимосвязанное и плавное образование всех элементов формирующегося нового типа организации, в данном случае птиц, а мозаичная эволюция, т. е. смешение старых и новых признаков.
Заключение Синтетическая теория эволюции на протяжении многих последних лет играла ведущую роль в развитии биологических исследований и привела к глубокому пониманию роли генетических факторов в эволюционном процессе. В настоящее время все большую роль в эволюционных исследованиях начинает играть экология. Благодаря появлению и развитию таких дисциплин как поведенческая экология, экоморфология, экология сообществ изучение экологических факторов эволюции вышло на принципиально новый уровень и ведет к лучшему пониманию сущности вида и механизмов видообразования.
Основные положения В отличие от синтетической, эпигенетическая теория видит эволюционный процесс прежде всего как процесс эволюционного преобразования онт
|
|||||||||||
|
|
Последнее изменение этой страницы: 2016-08-15; просмотров: 1404; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.141 (0.021 с.) |








 Дивергенция. Наиболее часто в ходе эволюции мы наблюдаем дивергенцию или расхождение признаков у видов, происходящих от общего предка. Дивергенция начинается на популяционном уровне, Она обусловлена различиями в условиях среды, в которых обитают и к которым по-разному приспосабливаются под действием естественного отбора дочерние виды. Определенную роль в дивергенции играет и дрейф генов. Дивергенция обусловливает увеличение числа видов и продолжается на уровне надвидовых таксонов. Именно дивергентной эволюцией обусловлено поразительное разнообразие живых существ.
Дивергенция. Наиболее часто в ходе эволюции мы наблюдаем дивергенцию или расхождение признаков у видов, происходящих от общего предка. Дивергенция начинается на популяционном уровне, Она обусловлена различиями в условиях среды, в которых обитают и к которым по-разному приспосабливаются под действием естественного отбора дочерние виды. Определенную роль в дивергенции играет и дрейф генов. Дивергенция обусловливает увеличение числа видов и продолжается на уровне надвидовых таксонов. Именно дивергентной эволюцией обусловлено поразительное разнообразие живых существ.





