Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Изолированная анемия сосудодвигательного центра при сохранности импульсов на него со стороны синокаротидной зоныСодержание книги
Поиск на нашем сайте
В настоящее время существуют две совершенно противоположные точки зрения о факторах, вызывающих повышение функциональной деятельности сосудодвигательного центра. Согласно одной точке зрения, сосудодвигательный центр не реагиру ет на изменения давления в сосудах самого центра, единственным регулятором кровяного давления является рецепторная синокаротидная зона [Букерт и Гейманс (Buckaert, Heymans), 1939; Геринг (Hering)]. По мнению Букерта, Гейманса и Геринга, низкое кровяное давление и уменьшение тока крови в сосудах мозга, вызванные выключением питающих мозг общих сонных или позвоночных артерий, не стимулируют непосредственно сосудодвигательный центр. Но уменьшение кровяного давления в каротидном синусе возбуждает располагающиеся там ре-цепторные окончания, благодаря чему осуществляется рефлекторное возбуждение сосудодвигательного центра через синусный нерв. В случае высокого кровяного давления в оинокаротидной зоне тот же нерв является проводником импульсов, угнетающих функциональную деятельность еосудодвигательного центра. В результате этого кровяное давление в мозговых сосудах рефлекторно снижается. Однако, согласно представлениям ряда исследователей, наряду с рецепторным аппаратом в синокаротидной зоне, регулирующим кровяное давление посредством влияний, идущих по синусному нерву, сам сосудодвигательный центр обладает непосредственной чувствительностью к понижению давления крови в сосудах ретикулярной субстанции продолговатого мозга. Работы, трактующие о подобной возможности, вышли главным образом из лаборатории Н. Н. Аничкова (П. П. Гончаров и И. Р. Петров, 1934; П. Н. Веселкин, 1937; К. К. Леонов, 1936—1937; И. Р. Петров, 1949). Эти авторы полагают, что понижение давления в сосудах продолговатого мозга является стимулом для повышения активности клеток еосудодвигательного центра. Экспериментальные доказательства высказанного положения были получены в лаборатории Н. Н. Аничкова при анемиза-ции мозга после предварительного выключения влияния на сосудодвигательный центр со стороны синокаротидной зоны. Опыт ставился следующим образом: выключались оба депрессорные и оба синокарогидные нервы, вследствие чего кровяное давление устанавливалось на уровне, значительно выше исходного. На фоне этого давления вызывалась анемия мозга закрытием обеих сонных и позвоночных артерий. Давление поднималось несколько выше. На основании результатов такого рода опытов эти исследователи заключили, что понижение давления в сосудах продолговатого мозга ведет к повышению деятельности еосудодвигательного центра. Однако какова природа раздражителя при анемии продолговатого мозга, этими исследованиями установлено не было. Иначе говоря, не было решено, возбуждается ли сосудодвигательный центр механическим понижением давления в сосудах или повышение тонического состояния его является следствием изменения химизма среды, окружающей клетки центра. Опыты, поставленные в лаборатории Н. Н. Аничкова с целью установить чувствительность еосудодвигательного центра к повышению давления в сосудах продолговатого мозга, положительных результатов не дали. Выяснилось, что такой чувствительностью указанный центр не обладает.
Диаметрально противоположные выводы в двух сериях исследований о регуляции деятельности еосудодвигательного центра заставляют обратить особое внимание на технику экспериментов, поставленных для решения столь важного вопроса. Выше мы уже видели, что Букерт и Гейманс пришли к выводу об отсутствии реакций со стороны дыхательного и еосудодвигательного центров в ответ на изменение давления в сосудах продолговатого мозга (в сосудах виллизиева круга). Такого рода мнение сложилось на основании опытов, в которых понижение давления в сосудах виллизиева круга от 80 до 40 мм ртутного столба не сопровождалось какими-либо нарушениями дыхательной деятельности или кровяного давления. Постановка эксперимента была весьма сложной. На одной стороне перевязывались все ветви каротидного синуса, за исключением внутренней сонной артерии. В общую сонную артерию той же стороны вводилась канюля, соединенная с ртутным манометром. Благодаря тому, что внутренняя сонная артерия оставалась свободной, возможна была регистрация кровяного давления в виллизиевом кругу. Синусный нерв перерезался. На противоположной стороне перевязывались все ветви каротидного синуса, но сохранялся синусный нерв. Таким образом, кровоснабжение всего мозга осуществлялось теперь только позвоночными артериями, а сосудо-двигательный центр испытывал нервное влияние со стороны только одной синокаротидной рецепторной зоны (рис. 104).
В этих условиях зажатие обеих позвоночных артерий вызывало падение давления в сосудах виллизиева круга, но дыхание и кровяное давление оставались без изменений. Несмотря на быстрое повышение давления в сосудах мозга при освобождении позвоночных артерий от зажимов, никаких изменений кровяного давления и дыхания не отмечалось. При рассмотрении хода эксперимента можно отметить целый ряд методических ошибок. Уже с самого начала опыта мозг ставится в условия пониженного кровоснабжения вследствие выключения основных питающих его артерий. Поэтому еще до зажатия позвоночных артерий мозговая ткань не только полушарий, но и продолговатого мозга испытывает некоторое кислородное голодание. Наложение зажимов на позвоночные артерии усугубляет общую анемию всего мозга в целом, так как, благодаря этой манипуляции, резко понижается давление не только в сосудах продолговатого мозга, но и в сосудах других его отделов. Кроме того, нарушается нормальная рефлекторная регуляция деятельности сосудодвигательного центра, благодаря перерезке одного из синусных нервов. Иначе говоря, результаты экспериментов Букерта и Гей-манса не исключают возможности регуляции функциональной деятельности вазомоторного центра с помощью изменений давления в сосудах продолговатого мозга. В экспериментах Букерта и Гейманса сосудодвигательный центр продолговатого мозга находится под постоянным влиянием со стороны одной синокаротидной зоны, посылавшей импульсы по сохраненному синусному нерву. При такого рода постановке опытов оставался открытым вопрос о центральной регуляции деятельности сосудодвигательного и дыхательного центра. Не вполне безупречны с методической стороны опыты и тех экспериментаторов, которые утверждают существование центральной регуляции сосудодвигательного и дыхательного центров наряду с рефлекторной регуляцией их деятельности. Некоторые из последователей этой точки зрения считают даже, что уменьшение давления в сосудах вазомоторного центра вызывает более мощную прессорную реакцию, чем влияния со стороны синокаротидной зоны (И. Р. Петров, 1949). Погрешности опытов исследователей данного направления состоят в том, что подобно Букерту и Геймансу изучение реакций сосудодвигательного центра на падение давления в сосудах, снабжающих его кровью, производится в условиях анемии всего мозга в целом. Исследование ведется на центрах продолговатого мозга, освобожденных от мощных, естественных стимуляторов. Перерезка синусных, а в некоторых случаях и аортальных нервов, кокаинизация или резекция бифуркации ведут к уничтожению рефлекторных тормозных влияний на данные центры и осо бому функционированию их, не имеющему места в нормальных условиях. Для того чтобы решить вопрос о характере реакций центров продол-говатого мозга в условиях анемизации его, необходима такая постановка опыта, при которой действию анемии подвергался бы только один про-долговатый мозг. Основным требованием при этом является сохранение нормального кровообращения в полушариях головного мозга и нормальной циркуляции крови через каротидный синус. Нормальная деятельность рецепторного аппарата синокаротидной зоны необходима потому, что, как мы уже видели, каротидный синус осуществляет постоянное тормозящее воздействие на сосудодвигательный центр продолговатого нозга.
Современная нейрохирургическая техника позволила нам поставить именно такой эксперимент. Опыты производились на собаках. Основанием для этого послужил тот факт, что преобладающее количество опытов, поставленных для выяснения функции синокаротидной зоны, было проведено на собаках. У собаки сосуды шеи и головы со всеми их ветвями и анастомозами изучены тщательнее и подробнее, чем у других лабораторных животных. Кроме того, величина головы собаки позволяет осуществлять подход к артериям на основании мозга и закрывать их. Известно, что вызвать анемию головного мозга собаки не легко. Многочисленные исследователи, в том числе А. П. Любомудров, М. Андреев, В. В. Колесников и другие, показали, что перевязка сонных и позвоночных артерий на шее собаки не ведет к полному обескровливанию мозга. Выключение сосудов, доставляющих кровь к мозгу в нормальных условиях, сопровождается расширением коллатеральных сосудов в той или иной мере возмещающих недостаточное питане головного мозга. В свое время Дэвис и Поллок (Davis a. Pollock, 1926) предложили так называемый «анемический метод» децеребрации кошек. Метод заключался в том, что при операции через рот производилась трепанация основной кости, благодаря чему основная артерия становилась доступной для перевязки. При одновременной перевязке обеих общих сонных артерий на шее кошки наступала децеребрационная ригидность, что указывало на анемию полушарий головного мозга и среднего мозга вместе с красными ядрами. При повторении опытов Дэвиса и Поллока мы отметили резкое повышение кровяного давления вследствие выключения синокаротидных зон при перевязке сонных артерий и падение температуры мозга до 34°. При изучении сосудистой сети мягкой мозговой оболочки через окно, ввинченное в череп, при таком способе достижения анемии отмечалось все же движение крови по артериям, хотя скорость тока крови и была незначительной. Позже метод Дэвиса был использован Уайтом (White. 1937), который перевязывал сонные артерии на шее и накладывал сере-брянный зажим (клипс) на основную артерию. Однако при такой постановке опыта анемия мозга не развивалась; децеребрационная ригидность появлялась только после дополнительной перевязки всех ветвей каротид-ного синуса.
Существуют и другие способы анемизации мозга собаки, например, метод Кэбат (Kabat, 1938) или метод, предложенный Э. А. Асратяном (1949). Ни один из указанных методов не мог быть использован для наших опытов, так как все они имели целью достижение анемии всего мозга в целом, в то время как нам нужно было добиться изолированной анемии одного продолговатого мозга. Иначе говоря, предстояло разработать способ, с помощью которого можно было бы вызвать анемию продолговатого мозга. Разработанный и использованный нами в опытах метод заключался в следующем. У взрослой молодой собаки под морфинно-эфирным наркозом с добавлением в местах разреза обезболивающих веществ обнажалась височная область мозга, достигалось это косым вертикальным разрезом кожи и височной мышцы, скусыванием скуловой дуги и коронарного отростка, а также подвисочной трепанацией и вскрытием твердой
мозговой оболочки. На втором этапе операции производился срединный разрез кожи дорзальной части шеи протяженностью от затылочного отверстия до III шейного позвонка. Снимались дужки I и II позвонка и обнажался спинной мозг. При операции использовался электронож и воск для того, чтобы в наибольшей степени избежать потери крови. После того как заканчивались все подготовительные хирургические процедуры, собака лежала в течение 20—25 минут совершенно спокойно. Затем ртутным манометром исследовалось общее кровяное давление в бедренной артерии до и после зажатия сонных артерий на шее.
На третьем этапе операции производилось закрытие артерий, по которым возможно поступление крови к центрам продолговатого мозга. Артерии зажимались маленькими серебряными зажимами (клипсами). Вначале закрывались задние соединительные артерии. Для этой цели голова собаки, поворачивалась на бок, шпателем осторожно приподнималась височная доля мозга, спинномозговая жидкость тщательно удалялась. После того как на основании мозга отчетливо выступали все образования, накладывался клипс на заднюю соединительную артерию между отходящими от нее задней мозговой и верхней мозжечковой артерией (рис. 109, /). Затем эта же манипуляция производилась и на другой стороне. В зависимости от расположения основной артерии на варо-лиевом мосту, закрытие этой арте-
рии производилось одновременно с наложением клипса на правую или левую заднюю соединительную артерию (рис. 109, 2). Таким образом, выключение указанных артерий разобщало кровоснабжение полушарий головного мозга от мозжечка, варолиева моста и продолговатого мозга. Полушария головного мозга и подкорковые узлы получали кровь нормальным образом по внутренним сонным артериям. Продолговатый мозг, варолиев мост и мозжечок снабжались системой позвоночной артерии. Основной анастомоз между системами внутренней сонной и позвоночной артерии в этих случаях был представлен затылоч-ной артерией на шее (рис. 199, А. осс.). После наложения клипсов на задние соединительные и основную артерию височные мышцы с обеих сторон сшивались для того, чтобы предохранить мозг во время дальнейшего опыта от каких-либо внешних воздействий (охлаждения, высыхания и т. д.). Затем временно зажимались обе общие сонные артерии на шее. Это давало возможность судить о силе реакций со стороны рецепторного аппарата каротидного синуса в условиях раздельного кровоснабжения полушарий и заднего отдела головного мозга. На следующем этапе опыта вскрывалась твердая мозговая оболочка над верхней частью спинного мозга и нижней частью продолговатого мозга и накладывались клипсы на обе верхние ветви позвоночной артерии (рис. 109, 3). Снова временно закрывались обе общие сонные арте-рииили обе сонные и обе позвоночные артерии на шее для определения степени тормозящего влияния синокаротидных зон на сосудодвигатель-ный центр продолговатого мозга. После выключения верхних ветвей позвоночной артерии (рис. 109, R. s. a. v.) кровоснабжение продолговатого мозга осуществлялось только по передней артерии спинного мозга (рис. 109, a, v, s). При наложении на нее клипса (рис. 109, 4) создавалась полная анемия продолговатого мозга. Кровь могла поступать в продолговатый мозг только по внутримозговым капиллярам, а также по анастомозам в мягкой мозговой оболочке как со стороны среднего мозга, так и со стороны спинного мозга. Но в соответствии с закономерностями коллатерального кровообращения в мозгу, установленным в нашей лаборатории, можно было полагать, что оба указанных способа компенсации далеко не могли возместить кровоснабжения продолговатого мозга, имеющего место в норме. Наши наблюдения указывают, что в некоторых случаях при выключении сосудов, снабжающих продолговатый мозг, заполнение артериальной сети его сосудов происходит за счет затекания крови из вен. Таким образом, разработанный нами метод выключения сосудов, питающих продолговатый мозг, создает наибольшую его анемизацию, доступную в опыте. В ряде опытов закрытию передней артерии спинного мозга предшествовало зажатие позвоночных артерий на шее собаки. В результате этого возникала хотя и значительная, но неполная анемия продолговатого мозга, так как кровь поступала к нему по анастомозам из артерий спинного мозга в переднюю артерию спинного мозга. С целью вариации степени анемии продолговатого мозга в некоторых экспериментах вместо передней артерии спинного мозга закрывались клипсами обе нижние ветви позвоночной артерии (рис. 109, R. I. a. v.). Опыты, проделанные нами совместно с Е. Н. Космарской (1949), показали, что при закрытии артерий в участках, показанных на рис. 109, возникает наиболее полная, доступная в опыте анемия продолговатого мозга. Но в зависимости от высоты кровяного давления и развивающегося впоследствии кровоснабжения по коллатеральным сосудам, выключение основных артерий, снабжающих продолговатый мозг, или ведет к полной его анемии или создает в нем анемию значительной степени. Соответственно тому, развивается ли в продолговатом мозгу полная анемия или она достигает критической степени (являясь не совсем
Рис. 110. Изменение кровяного давления и дыхания при полной анемии продолговатого мозга. Стрелкой указан момент наложения клипса на переднюю артерию спинного мозга, последнюю артерию из снабжавших продолговатый мозг. полной вследствие развития коллатерального кровоснабжения), можно констатировать несколько типов реакций со стороны дыхательного и сосудодвигательного центра. Описываемый ниже тип реакции характерен для случаев, в которых закрытие артерии ведет к полной анемии продолговатого мозга. Для иллюстрации приведем один из наших опытов. У собаки под морфинным наркозом кровяное давление, измеренное в бедренной артерии, равнялось 100 мм ртутного столба. После наложения клипсов на задние соединительные артерии, основную артерию и верхние ветви позвоночной артерии кровяное давление и дыхание не изменились. Как можно видеть на рис. 110, закрытие передней артерии спинного мозга (последней из артерий, снабжавших продолговатый мозг) вызвало резкие нарушения деятельности дыхательного и сосудодвигательного центра, а также нарушение деятельности центра блуждающего нерва. Непосредственно после закрытия указанной артерии дыхание стало более редким, амплитуда дыхательных движений постепенно снижалась. Давление крови начало увеличиваться и на 40-й секунде после закрытия передней артерии спинного мозга кровяное давление поднялось на 14 мм ртутного столба. Одновременно с этим резко увеличилась пульсовая волна и частота сердечных сокращений, что указывало на прекращение тормозных влияний со стороны центра блуждающего нерва на сердечную деятельность. Последующие изменения дыхания и кровяного давления говорят о патологическом функционировании центров продолговатого мозга в условиях его анемизации. Такая резко измененная деятельность продолжалась всего 4 минуты после полного прекращения притока крови к продолговатому мозгу, затем последовала смерть собаки от остановки дыхательной деятельности. Обращает на себя внимание, что кровяное давление начало прогрессивно снижаться, прежде чем прекратилось дыхание. Инъекция сосудов мозга пластмассой показала, что анемия продолговатого мозга была полной, так как сосуды его пластмассой не заполнялись (рис. 111). Употреблявшаяся нами пластмасса была крупно дисперсной и заполняла все артерии до капилляров. Таким образом, в этом и аналогичных ему опытах полная анемия продолговатого мозга вела к кратковременному и незначительному повышению общего кровяного давления. Однако наступавшие одновременно с этим изменения дыхательной и сердечной деятельности отчетливо указывали на патологическую деятельность центров продолговатого мозга, функционировавших только до того момента, пока не истощались энергетические возможности нервных клеток этих центров. Через 4—7 минут после полного прекращения поступления крови к клеткам дыхательного и сосудодвигательного центра животное погибало от остановки дыхательной и сердечной деятельности. Следовательно, наступающее в таких опытах повышение кровяного давления нельзя рассматривать как нормальную реакцию сосудодвигательного центра на полную анемию, регулирующую его деятельность. Напротив, повышение кровяного давления представляет собой проявление патологической, агональной деятельности нервных клеток сосудодвигательного центра. Таким образом, результаты наших опытов указывают на то, что при полной анемии продолговатого мозга у собак имеет место гибель нервных клеток сосудодвигательного и других расположенных в нем центров. Другими словами, в этих случаях с особой отчетливостью выступает патологическое действие анемии на сосудодвигательный центр.
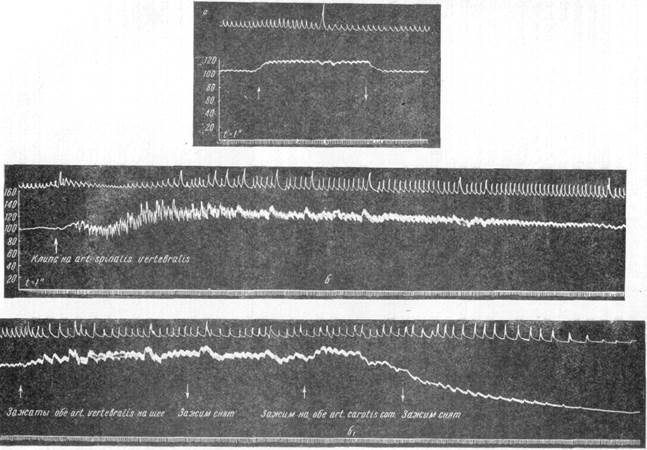
Рис. 111. Мозг собаки, сосудистая система которого была налита пластмассой, после опыта анемии продолговатого мозга (инъекция в общую сонную артерию). Обращают на себя внимание заполнение пластмассой сосудов полушарий головного мозга и отсутствие ее в сосудах варолиева моста, м'озжечка, продолговатого мозга и верхней части спинного мозга. На схеме в правом нижнем углу показаны места, где на артерии накладывались клипсы. Рассмотрим теперь те случаи, когда закрытие артерий, снабжающих продолговатый мозг, имело своим следствием развитие анемии критического уровня. Анемия в указанных случаях была неполной, так как коллатеральное кровоснабжение продолговатого мозга развивалось в значительно большей степени, чем это имело место в ранее рассмотренной группе опытов. Более интенсивный приток крови к центрам продолговатого мозга обнаруживался при инъекции пластмассы в сосудистую сеть мозга после смерти животного. Если при полной анемии никогда не наблюдалось прохождения инъекционных масс даже в сосуды мозжечка, то при критической, но не полной анемии пластмассу можно было обнаружить в сосудах верхней части полушарий мозжечка, иногда в сосудах моста или даже в сосудах верхних сегментов спинного мозга. Поступление хотя и незначительных количеств крови к дыхательному и со-судодвигательному центру продолговатого мозга являлось причиной особенностей их поведения в условиях сохранения постоянных воздействий со стороны синокаротидной зоны. В зависимости от особенностей реакций данных центров на различную степень анемии, все опыты с критической анемией продолговатого мозга могут быть разбиты на три группы. Таким образом, в то время как в условиях полной анемии продолговатого мозга наблюдается лишь один тип реакции дыхательного и сосудодвигательного центра, при критической анемии можно выделить три различных типа реакций указанных центров. Рассмотрим каждый из этих трех типов реакций. В первой группе опытов с критической анемией выключение всех артерий, снабжающих продолговатый мозг кровью, не влекло за собой вообще никаких изменений кровяного давления и дыхания. На записях кровяного давления и дыхания можно проследить, что последовательное закрытие задних соединительных артерий, основной артерии, верхних ветвей позвоночной артерии и передней артерии спинного мозга никак не сказалось на величине общего кровяного давления. Кровяное давление осталось на том же уровне, который оно имело в начале опыта (рис. 112, а, б, в). Выше мы уже указывали, что деятельность сосудодвигательного центра находится в тесной зависимости от постоянных рефлекторных влияний со стороны рецепторных синокаротидных зон. Данные современной электрофизиологии позволяют говорить о тормозном характере импульсов, поступающих от синокаротидных зон к клеткам сосудодвигательного центра. Сохранение рефлекторных воздействий синокаротидных зон на клетки сосудодвигательного центра являлось одним из необходимых условий при постановке наших опытов. Изучение характера реакций рецепторного аппарата каротидного1 синуса при понижении давления в нем позволило отметить, что при наличии нормальных рефлекторных влияний со стороны синокаротидных зон на сосудодвигательный центр анемия его не ведет к повышению кровяного давления (рис. 112, б). Но если в том же опыте устранить рефлексы с синокаротидных зон кокаинизацией синусного нерва или бифуркации, кровяное давление повышается (рис. 112, в). Во второй группе опытов повышение кровяного давления отмечалось в тех случаях, когда анемия продолговатого мозга достигала критического уровня, а тормозящие влияния со стороны синокаротидных зон были слабо выражены. В экспериментах этой группы зажатие обеих сонных артерий на шее собаки вызывало повышение общего кровяного давления всего на 8—20 мм ртутного столба (рис. 113, а). Это обстоятельство указывало на незначительность тормозных влияний со стороны
в Рис. 112. Первый вид деятельности сосудодвигательного и дыхательного центра в условиях критической неполной анемии. а - повышение давления при зажатии сонных артерий на шее до опыта; б—давление осталось на исходном уровне после к артерии, снабжающих продолговатый мозг; в —повышение давления при новокаинизации каротидного синуса. синокаротидной зоны на деятельность сосудодвигательиого центра. В таких условиях закрытие последней артерии, снабжающей продолговатый мозг кровью (в наших опытах — передней артерии спинного мозга), вызывало повышение общего кровяного давления. Иллюстрирующие данное положение записи одного из опытов позволяют проследить все изменения дыхания и кровяного давления с момента критической неполной анемизации продолговатого мозга до момента смерти животного (рис. 113, б). Сравнение этих записей с записями опытов, в которых у собак была вызвана полная анемия продолговатого мозга, позволяет отметить, что характер реакций дыхательного и сосудодвигательного центра как в том, так и в другом случае один и тот же. Различие между ними состоит только в том, что при критической анемии продолговатого мозга процессы больше растянуты во времени. В обоих случаях повышение общего кровяного давления представляет собой патологическую реакцию сосудодвигательного центра в ответ на лишение его нормального кровоснабжения. Следовательно, и в случаях критической неполной анемии она не может рассматриваться как фактор, регулирующий жизнедеятельность сосудодвигательного центра в нормальных условиях. Повышение кровяного давления, вызванное критическим уровнем анемии продолговатого мозга, является результатом патологических сдвигов в клетках сосудодвигательного центра, погибающих в течение того или иного срока в зависимости от возможности поступления к ним крови по коллатеральным сосудам. В третьей группе опытов наблюдаются несколько иные соотношения, так как в этих случаях при наличии критической неполной анемии отмечается значительное тормозное влияние синокаротидной рефлекторной зоны на сосудодвигательный центр. В этих случаях (рис. 114, а) устранение тормозных импульсов при зажатии сонных артерий на шее и падении давления в синусе ведет к значительному повышению кровяного давления. Таким образом, синокаротидные рефлексогенные зоны оказывают у этих животных большое по силе воздействие на деятельность сосудодвигательного центра в продолговатом мозгу. Если у собак с такого рода сильным рефлекторным влиянием каро-тидного синуса производилась полная анемизация продолговатого мозга, то наступали изменения дыхательной и сердечной деятельности, подобные изменениям, представленным на рис. 114, б. При рассмотрении его можно сделать заключение, что кровяное давление испытывает в этих случаях значительные колебания в результате одновременного воздействия на сосудодвигательный центр сильных тормозных импульсов со стороны каротидного синуса и анемизации всех центров продолговатого мозга. Эффект, связанный с изменением тормозного влияния синокаротидных зон, отчетливо выступает при выключении их зажатием обеих сонных артерий. Наступающее при этом повышение кровяного давления указывает, какого уровня оно могло бы достигнуть при критической анемии, если бы были устранены рефлекторные влияния со стороны синокаротидной зоны. В экспериментах такого рода с очевидностью обнаруживается значение рефлекторной регуляции кровяного давления. Даже в условиях нарушения нормальных процессов жизнедеятельности нервных клеток сосудодвигательного центра тормозной синокаротидный рефлекс препятствует проявлению патологической деятельности их, способствуя удержанию кровяного давления на уровне, характерном для каждого отдельного животного.
Рис. 113. Второй вид деятельности сосудодвигательного и дыхательного центра в условиях критической неполной анемии, а — изменение кровяного давления при зажатии обеих общих сонных артерий на шее до опыта; б и б1 — кровяное давление и дыхание после прекращения притока крови к продолговатому мозгу по всем снабжающим его артериям. Таким образом, при одновременном действии на сосудодвигательный центр анемии и рефлекторных влияний с синокаротидных зон тормозная импульсация, поступающая по синусному нерву, оказывается более эффективной, чем действие анемии. Лишь при исключении рефлекторных воздействий проявляется патологическая активность нервных клеток сосуд одвигательного центра, процессы жизнедеятельности в которых нарушены анемией. Можно думать, что тормозные импульсы, идущие со стороны каротидных синусов в нормальных условиях жизнедеятельности клеток сосудодвигательного центра, тормозят протекающие в них процессы обмена веществ. Устранение тормозных влияний перерезкой синусных нервов или кокаинизацией их или бифуркаций сонных артерий ведет к проявлению всех функциональных возможностей клеток указанного центра, в результате чего кровяное давление повышается. Следует полагать, что при анемии недостаток кислорода и питательных веществ ведет к усиленной трате всех энергетических запасов, имеющихся в нервной клетке. Прекращение выведения продуктов обмена веществ вызывает отравление нервной клетки. В таких случаях наличие тормозного влияния со стороны синокаротидных зон, понижающего процессы обмена веществ в нервных клетках сосудодвигательного центра, способствует более экономному расходованию их энергетических ресурсов. Это в свою очередь уменьшает интенсивность процесса накопления продуктов обмена веществ, и таким образом клетка отравляется более медленно. В связи с этим представляют интерес данные Бронка (1938), отметившего, что анемия значительной степени, вызывающая разрушение молекулярной структуры клеток дыхательного и сердечно-сосудистого центра, сопровождается спонтанными разрядами. В соответствие с этими данными можно поставить повышение кровяного давления, обычно наблюдающееся в случаях смерти животного через 4—7 минут после выключения всех артерий, снабжающих продолговатый мозг. Возможно предположить, что в этих случаях слабое тормозное влияние со стороны каротидного синуса ведет к быстрому расходованию энергетических ресурсов нервных клеток сосудодвигательного центра, причем распад их структуры сопровождается значительным увеличением функциональной деятельности. Увеличение функциональной деятельности находит свое отражение в повышении кровяного давления. Кроме того, полная анемия приводит к нарушению деятельности дыхательного центра и центра блуждающего нерва, расположенных в продолговатом мозгу. Увеличение содержания углекислоты в крови и ослабление тормозных влияний блуждающего нерва на сердечнососудистую деятельность в свою очередь должно способствовать повышению уровня кровяного давления. В тех же случаях, когда синокаротидные рецепторные зоны оказывают непрерывное нормальное тормозное влияние на сосудодвигатель-ный центр, распад в его клетках идет более медленно. Анемия ведет к постепенному затиханию процессов жизнедеятельности нервных клеток дыхательного, сосудодвигательного и других центров продолговатого мозга, в результате чего животное погибает в условиях постепенного понижения общего кровяного давления и ухудшения дыхательной деятельности. Полученные в наших опытах данные делают сомнительным существующее в литературе мнение, согласно которому наиболее чувствительными к аноксии в центральной нервной системе являются синапсы. Приведенные опыты (рис. 114) свидетельствуют о том, что при полной 217-
Рис. 114. Третий вид деятельности сосудодвигательного и дыхательного центра в условиях критической неполной анемии, а — кровяное давление и дыхание при зажатии обеих общих сонных артерий на шее; б — кровяное давление и дыхание после закрытия передней артерии спинного мозга, последней из артерий, снабжавших его кровью стрелками показан момент наложения клипса на названную артерию, а также наложение и снятие зажимов с общих сонных артерий па шее анемии синаптические связи между разветвлениями синусного нерва и клетками дыхательного и сосудодвигательного центра продолговатого мозга не прерываются.
Рис. 115. Выключение артерий в опытах с полной анемизаци- ей всего головного мозга. Обозначения на схеме те же, что и на рис. 109. При изучении деятельности сосудодвигательного центра в условиях анемии мы предприняли ряд опытов, в которых реакции указанного центра наблюдались при полной анемии всего мозга в целом. Для осуществления анемии всего мозга мы закрывали обе внутренние сонные артерии после вхождения их в полость черепа на основании мозга,
Рис. 116. Реакции сосудодвигательного и дыхательного центра в опыте с полной анемией всего головного мозга в целом. а —кровяное давление и дыхание в начале опыта; б —кровяное давление и дыхание после клипирования обеих внутренних сонных артерий; в — изменение кровяного давления и дыхания во время полной анемии всего головного мозга. накладывали клипсы на обе верхние ветви позвоночной артерии и пе-реднюю артерию спинного мозга так, как это показано на рис. 115. Результаты, полученные нами в серии опытов такого рода, дали возможность констатировать, что полная анемия мозга при сохранении реф-лексов с каротидных синусов также не ведет к повышению общего кро-вяного давления. Для иллюстрации сказанного выше приводим запись реакций сосудодвигательного и дыхательного центра, взятую из одного опыта, типичного для всей серии. При рассмотрении рис. 116, а, б, в можно отметить, что последовательное выключение артерий, снабжающих головной мозг, ведет к постепенному снижению деятельности центров продолговатого мозга. Полная анемия всего мозга в целом, так же как и полная анемия одного продолговатого мозга, приводит животное к смерти через 6—7 минут после выключения последней артерии. Подводя итог краткому изложению полученного нами экспериментального материала, следует указать, что изучение влияния анемии на деятельность сосудодвигательного центра представляет собой сложную задачу. Прежде всего чрезвычайно сложной является методическая сторона эксперимента. Анемия продолговатого мозга требует выключения наибольшего числа артерий, снабжающих его кровью. Но даже в усло-виях прекращения притока крови по всем артериям остается возможность кровоснабжения продолговатого мозга по коллатеральным со-судам. Анатомически коллатеральными сосудами, различно выраженными у различных животных, может служить капиллярная сеть мозгового вещества, а также хорошо развитая сосудиста
|
||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-01; просмотров: 195; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.147.45.159 (0.02 с.) |