Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Морфологічно-диференційовані структури прокаріотСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
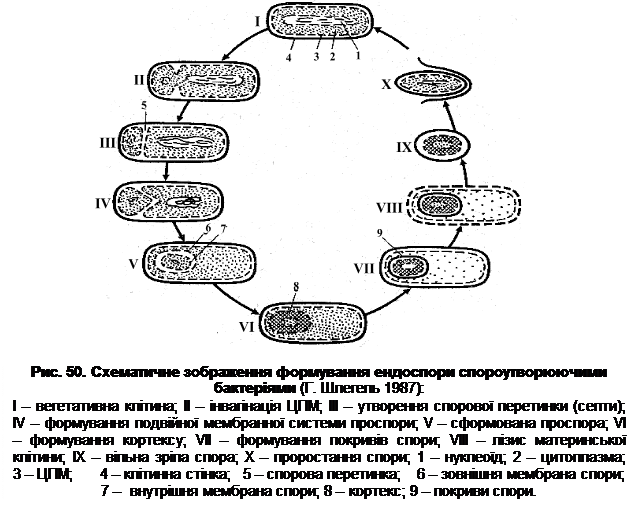
Ендоспора – це стадія спокою бактеріальної клітини, яка сприяє виживанню (збереженню) бактерій в несприятливих умовах середовища, тобто ендоспора не служить для розмноження. Вважається, що утворення ендоспори є однією з фаз розвитку мікробної популяції. Але ця стадія не є обов’язковою для життєвого циклу спороутворюючих бактерій. За сприятливих умов живлення клітини можуть необмежений час розмножуватися, не переходячи в стадію спокою. Процес спороутворення можна також інгібувати, наприклад, внесенням у середовище глюкози або частими пересівами культури. Роль інгібуючого фактора залежить від часу його внесення. Так, наприклад якщо до суспензії вегетативних клітин B. mycoides протягом перших 5 год після їх внесення у воду додати глюкозу, то утворення спор репресується. Глюкоза, внесена пізніше як через 6 год, уже не діє – інгібування спороутворення не відбувається. В той же час індукція (дерепресія) спороутворення зберігається в період між 10 і 13-ю годинами після внесення бацил у воду – приблизно 90% клітин утворюють спори. Таким чином, процес споруляції певною мірою регулюється факторами середовища. Процес споруляції можна індукувати. До факторів, що індукують утворення ендоспор, відносять: нестачу поживних речовин, надлишок продуктів метаболізму, наявність у середовищі іонів Mn2+, Ca2+, зміну температури вирощування, рівня pH, концентрації О2 та ін. Отже, спороутворення відбувається у тих випадках, коли загальні умови існування допускають його. Якщо помістити вегетативні клітини на певній стадії розвитку в дистильовану воду, то можна спостерігати так звану ендотрофну споруляцію, тобто утворення спор за рахунок запасних речовин клітини. В таких випадках утворення спор дійсновикликається дефіцитом екзогенного субстрату. Тривалість процесу споруляції в різних представників ендоспороутворюючих бактерій різна. В аеробних мезофільних бацил, наприклад, спороутворення займає близько 8 год. Здатність клітин до утворення спори визначається комплексом генів, який називають спорулоном, хоч він і не є цілісно оформленою структурою. Проте встановлена наявність кількох груп – оперонів, локалізованих у різних ділянках хромосоми. Було показано, що B. subtilis має 42 спорових оперони, кожен з яких включає ~3 гени. В цілому, B. subtilis має 150-200 генів, що контролюють процес споруляції. Події, які відбуваються на кожній стадії споруляції, можуть залежати від активності генів з різних оперонів. Найкраще процес спороутворення досліджено в представників родів Bacillus i Clostridium, хоча інформація отримана протягомостанніх років свідчить про принципову однотиповість цього процесу в усіх видів ендоспороутворюючих бактерій. Спороутворення проходить у декілька фаз (етапів). Зміна кожної стадії визначається активацією відповідних генів. При цьому один з продуктів гена, що “функціонує” індукує активність наступного гена. Для початку спороутворення необхідне завершення реплікації бактеріальної хромосоми. При цьому сигнал до спороутворення має надійти ще в період реплікації. Якщо реплікація хромосоми завершується в багатому на поживні речовини середовищі, тоді такий сигнал не надходить, а популяція продовжує вегетативний поділ. В процесі утворення спори хромосома не реплікується, і в спору переходить одна або декілька повністю реплікованих раніше хромосом. Клітина, що готується до споруляції, має не менше двох нуклеоідів, які утворюють тяж, який проходить майже по всій довжині клітини, включаючи в себе всю ДНК (рис. 50). У деяких видів роду Clostridium він утворюється лише частиною наявної в клітині ДНК і локалізується на одному з полюсів клітини, займаючи незначну частину її довжини. В подальшому від осьового тяжа відшнуровується ділянка, яка відповідає хромосомі. Вона переміщується до одного з полюсів клітини. Далі ЦПМ починає інвагінуватися, утворюються дві різного розміру структури, розділені мембраною. На цьому етапі процес споруляції є ще оборотним. Якщо бактеріальні клітини перенести в сприятливі для розвитку умови, то процес їхнього поділу завершиться утворенням двох вегетативних клітин різної довжини. Було показано, що цей процес контролюється генами восьми оперонів, локалізованих у різних ділянках хромосоми. На перших етапах споруляції використовуються запасні речовин і частина білків материнської клітини. У більшості видів роду Bacillus відбувається синтез специфічних протеаз, яким приписують різні функції, в тому числі участь у кругообігу білка, модифікації ДНК-залежних РНК-полімераз і білків-попередників покривів спори.
В подальшому ЦПМ більшої частини клітини починає оточувати меншу структуру, в результаті утворюється структура, обмежена двома ЦПМ, – проспора. Дві мембрани, які оточують проспору, розташовані одна щодо одної ніби в дзеркальному відображенні – зовнішній, раніше звернутий до середовища шар внутрішньої мембрани, повернутий на зовнішній бік, а в зовнішньої мембрани – всередину. В подальшому проспора відділяється від ЦПМ материнської клітини і ніби вільно “плаває” в ній. На цьому етапі процес споруляції стає необоротним, змінюються властивості мембрани, особливо її транспортні функції. Процес стає необоротним не лише для проспори, а і для материнської клітини, котра вже не може повернутися до вегетативного росту. Якщо така система потрапляє в несприятливі умови, вона або завершує утворення спори, або гине.
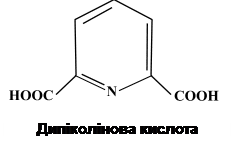
У проспорі посилюється поглинання іонів Са2+, розпочинається синтез деяких амінокислот та дипіколінової кислоти (піридин-2,6-дикарбонова кислота). Вегетативні клітини не здатні до синтезу останньої. Проспора, що утворилася, переміщується до центра клітини або залишається біля одного з її полюсів, що визначається видовою належністю об’єкта, тобто типом розташування спори. Між двома ЦПМ у більшості представників роду Bacillus утворюється ще одна оболонка – кортекс. Процес йде у дві стадії. Спочатку синтезується примордіальний (зачатковий) шар, який у процесі проростання спори дає оболонку проростка. Над примордіальним шаром синтезується модифікований шар муреїну, який і утворює кортекс. Синтез речовини кортексу продовжується до дозрівання спори. Цей біополімер є більш кислим, ніж його аналог у клітинній стінці. Пояснюється це тим, що багато залишків мурамової кислоти не містять пептидів, а значить – мають більше вільних карбоксильних груп. У B. cereus одночасно із синтезом кортекса формується екзоспоріум. На початку він синтезується на одному з полюсів проспори, а потім обростає її повністю. У більшості видів роду Clostridium початок закладки покривів передує формуванню кортекса. На зрізах спори кортекс має вигляд масивного електроно-прозорого шару. У спори, яка перебуває в стані спокою, він має гомогенний характер, а в спор, які проростають, – фібрилярний вигляд. Екзоспоріум нагадує мішок, в якому знаходиться спора. В різних видів будова екзоспоріума різна. Найбільш детально він вивчений у B. cereus. До складу екзоспоріума входять білки, ліпіди та вуглеводи. Вважається, що він захищає спору від дії зовнішніх факторів. Екзоспоріум C. botulinum має 15 паралельних шарів, загальна товщина яких більше 100 нм. У C. pasteurianum це тоненька структура, яка на одному з полюсів має велику пору. В деяких випадках зовнішній екзоспоріум покритий придатками або виростами різної форми. Наприкінці формування екзоспоріума молода спора ще осмотично нестабільна й нежиттєздатна, але в неї вже змінюються деякі фізичні властивості, наприклад, посилюється заломлення світла, що свідчить про синтез білкових сполук, оскільки в присутності інгібіторів синтезу білка світлозаломлення не підвищується. У подальшому продовжують формуватися оболонки спори. Із зовнішнього боку зовнішньої мембрани формується оболонка спори. Сама зовнішня мембрана на зрізах майже не визначається. У спорі продовжує накопичуватися дипіколінат кальцію, підвищується її стійкість до органічних розчинників, наприклад хлороформу. Спора набуває осмотичної стабільності, але ще не терморезистентна. Розвиток спори визначається активністю двох геномів – власного і материнського. Материнська частина клітини – спорангій – до останнього моменту зберігає свою активність. При цьому геном материнської клітини визначає синтез дипіколінової кислоти, білків, покривів спори (матеріалу), екзоспоріума та виростів спори. Цитоплазма спори оточена ЦПМ, яка переходить у примордіальний шар, а потім у кортекс. Речовина останнього становить 20-60% об’єму спори, але не перевищує 15% її маси. У процесі дозрівання спора зневоднюється. Вміст води у зрілій спорі знижується до 10-15%, що супроводжується інактивацією ферментів, а отже, гальмуванням метаболічних процесів. Спора набуває терморезистентності. Структура зрілої спори в різних ендоспороутворюючих бактерій майже однакова. В цитоплазмі знаходиться ядерний матеріал. Вміст білків і нуклеїнових кислот спори досягає 50-60% сухої маси. Значну частину спори становить дипіколінова кислота. В зрілої споривона знаходиться у формі солі – дипіколінату кальцію, вміст якого може становити до 10-15% за сухою масою. Кортекс обмежений зовнішньою мембраною спори, яка межує із зовнішньою оболонкою. Речовина останньої забезпечує стійкість спори до дії лізоциму. Спороутворення супроводжується активним синтезом білків, котрі на відміну від білків вегетативної клітини багаті на цистеїн та гідрофобні амінокислоти, з чим теж пов’язують стійкість спори до дії несприятливих факторів середовища. Оболонки спори забезпечують її стійкість до дії ферментів, проникності для органічних розчинників, поверхнево-активних речовин, антибіотиків, барвників тощо. Після того, як спора сформувалась, відбувається лізис клітинної стінки і спора звільняється. Схема будови зрілої спори представлена на рис. 51.
Спори мають специфічні властивості: по-перше, вони метаболічно неактивні; по-друге, вони стійкі до дії високих температур, хімічних речовин та опромінення. Природа (механізм) терморезистентності спори повністю не з’ясована. На думку Г. Шлегеля, це явище можна пояснити низьким вмістом води. Існує також думка, що терморезистентність приблизно пропорційна вмісту дипіколінової кислоти. Стійкість спор до дії хімічних речовин забезпечується також непроникністю зовнішніх покривів. Крім того, білки покривів спори захищають її від дії літичних ферментів. Спори різних бактерій відрізняються за формою, розмірами і локалізацією в материнській клітині (рис. 52). Так, у багатьох бактерій спора розташована в центрі клітини, а її діаметр не перевищує поперечника клітини – центральний (бацилярний) тип локалізації (B. subtilis, B. anthracis, B. megaterium). Спори багатьох бактерій мають овальну форму, але їх діаметр більший від поперечника клітини – клостридіальний тип локалізації. Клітина набуває при цьому вигляду веретена (C. pasteurianum). Спора може локалізуватися на одному з полюсів мікробної клітини, роздуваючи її спорангій. Сама клітина набуває форми барабанної палички – плектридіальний тип розташування спори (C. tetani). Спора займає центральну частину спорангія, але один його бік при цьомудеформується – латеральне положення (B. laterosporus).
Особливістю спороутворення в анаеробних бактерій є утворення на спорах особливих виростів різноманітної структури (рис. 53). Ця ознака специфічна, спадково закріплена і досить стійка. Навіть у дефектних спор, які втратили здатність утворювати оболонку, вирости зберігаються і не змінюють своєї структури. Вирости можуть мати форму тонких ниток або джгутиків, трубочок різної товщини, які розташовуються на всій поверхні спори або зібраних в пучок йоржикоподібних паличок, довгих широких стрічок, шипів, утворів, які нагадують роги оленя. Формування виростів розпочинається на ранніх стадіях споруляції – перед закладкою кортекса, потім вони ростуть, подовжуються, Цитоплазма материнської клітини навколо виростів лізується. На зрілих спорах, які вийшли із спорангія, вирости розпускаються.
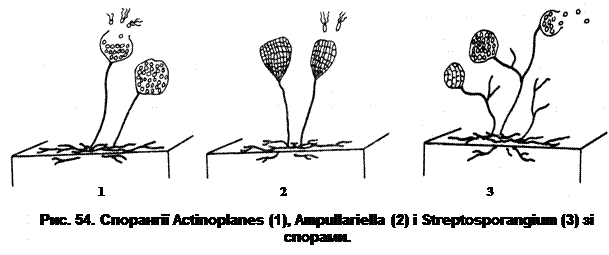
Речовина виростів складається переважно з білків, а також незначної кількості полісахаридів і ліпідів. Функції виростів вивчаються. Деякі автори вважають, що вирости на спорах – специфічно чутливі (хемосенсорні) органели, які подають „сигнал” на проростання спори у сприятливих для неї умовах. Інші вважають, що вирости виконують важливу функцію в процесі дозрівання спор, беруть участь у формуванні спорових покривів і кортексу. Існує також думка, що вирости на спорах – наслідок певних порушень обміну речовин. Процес проростання спори можна поділити на три етапи: ініціація, активація й виростання. Ендоспора навіть у сприятливих умовах може не проростати. Цей процес необхідно ініціювати. Активація означає готовність спори до проростання, при цьому зберігається її стійкість до температури, світлозаломлення та інші характеристики. Одним з ініціюючих факторів є термообробка. Було показано, що процес активації йде тим швидше чим вища температура. Наприклад, при 650С активація займає 45 хв, а при 340С для активації спори необхідно біля 48 год. Активація є процесом необоротним, вона триває декількахвилин і протікає у певній послідовності: спочатку знижується терморезистентність і відбувається вихід у середовище дипіколінату кальцію, після чого спора набуває здатності фарбуватися; далі спора втрачає діамінопімелінову кислоту і глюкозамін, а потім знижується світлорозсіювання. Процес ініціації залежить від температури, вологості середовища, pH та інших факторів. До початку проростання спора інтенсивно поглинає воду і набрякає. Протягом перших хвилин проростання вона втрачає біля 25% своїх білків, тобто тих білків, що синтезувалися в процесі споруляції. Гідроліз білків здійснюється за рахунок специфічної протеази самої спори, яка діє лише на ці білки. У ході проростання спори її оболонки лізуються або розриваються, а проросток виходить з оболонок. Виростання – це процес активного росту, в ході якого активно синтезуються РНК і білки, а через 1-2 год розпочинається реплікація ДНК. Спори актиноміцетів формуються переважно на повітряному міцелії (спороносцях), а в окремих випадках – на субстратному. Розташування спороносних гілок у деяких актиноміцетів – ознака досить закономірна й постійна. За формою найчастіше зустрічаються спіральні (з множинними завитками, примітивні спіралі, розтягнуті або у вигляді клубочків) спороносці. За характером розташування на спороносних гілках їх поділяють на мутовчастий, супротивний та послідовний типи. Утворення спор актиноміцетами може відбуватися різними способами. До найбільш розповсюджених та вивчених належить утворення спор шляхом фрагментації або сегментації спороносних гілок. Фрагментаційний процес характеризується тим, що навколо ядерних елементів, рівномірно розподілених по гіфі, концентрується цитоплазма, яка покривається власною оболонкою і перетворюється на зрілу спору. Оболонка спороносних ниток деякий час зберігається, але з часом розпадається, а спори висипаються і за відповідних умов знову проростають, утворюючи гіфи нових колоній актиноміцетів. При сегментаційному утворенні спор спостерігається простий поділ спороносних гілок. По всій довжині гілки утворюються поперечні перетинки, і спороносець ділиться на ряд рівномірних клітин-спор. Перед сегментацією в спороносній гілці відбувається поділ ядерної речовини – утворюється стільки зерен хроматину, скільки буде спор. Навколо хроматинових зерен також концентрується цитоплазма клітини. Коли спора дозріває, спороносець розпадається на окремі спори. У деяких родів актиноміцетів (Actinoplanes, Ampullariella, Streptosporangium та інші) утворюються спорангії-мішки, в яких містяться спори (рис. 54).
Окремі актиноміцети утворюють незвичні структури. Так, наприклад, представники родів Dermatophilus, Frankia, Geodermatophilus утворюють велику кількість спор у результаті поділу гіф в кількох площинах, а не перпендикулярно осі. Такі структури називають багатогніздими спорангіями. Деяким актиноміцетам властиве утворення склероцій – кулястих структур, які містять не спори, а клітини, заповнені ліпідами. Склероцій проростає як єдине ціле, утворюючи при цьому одну ростову трубку. У деяких актиноміцетів спорангіїв немає (наприклад, у представників родів Actinоbifida, Microbispora, Micromonospora, Micrоpolyspora). Поодинокі спори, їх пари або ланцюжки (до 20 спор), як форма розмноження та спокою, знаходяться безпосередньо на гіфах міцелію або на дуже коротких спороносних гіфах (спорофорах). Спори актиноміцетів відрізняються між собою за формою. Вони бувають кулясті, циліндричні, паличкоподібні, грушоподібні. Поверхня спор різних видів актиноміцетів теж може бути різною. На поверхні виявляються структури шипоподібної, горбкуватої, волосоподібної форми, в окремих випадках поверхня оболонки буває гладенька. Ця ознака використовується для визначення належності актиноміцетів до тієї чи іншої групи. Спори актиноміцетів більш стійкі до дії несприятливих умов довкілля, ніж вегетативні клітини. Так, культури актиноміцетів, висушені до повітряно-сухого стану, можуть зберігати життєздатність протягом кількох років. Спори стійкі також до різних механічних впливів: при розтиранні культур у суміші з частинками піску спори не втрачають своєї життєздатності, тимчасом як відповідні їм за розмірами фрагменти вегетативного міцелію гинуть. Проростання спор актиноміцетів однотипове. На свіжому поживному середовищі вони спочатку дещо набухають, потім на їх поверхні з’являються горбочки – бруньки, які, подовжуючись, проростають у довгі нитки. Нитки галузяться, формується міцелій – колонії актиноміцетів. Цисти, як форми спокою, виявляються в різних груп бактерій: азотобактера, спірохет, міксобактерій, рикетсій. При цьому в більшості міксобактерій утворення цист (міксоспор) – це одна зі стадій життєвого циклу. Після завершення стадії активного розмноження клітини міксобактерій групуються й утворюють так звані плодові тіла, котрі являють собою масу слизу, в який занурені клітини. Всередині плодових тіл клітини переходять у стан спокою (рис. 55). Утворення міксоспор супроводжується синтезом специфічного білка. ДНК не синтезується, а переходить з вегетативної клітини. Міксоспора може мати три-чотири копії хромосоми. Цисти міксоспор більш стійкі до нагрівання, висушування, дії фізичних факторів, ніж вегетативні клітини, тобто ця структура певною мірою, можерозглядатися як форма спокою.
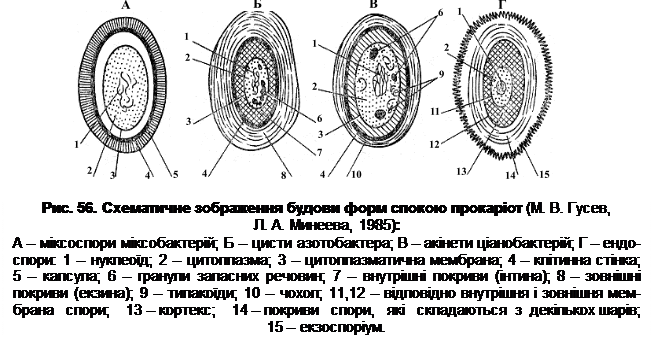
У деяких видів бактерій цисти морфологічно не відрізняються від вегетативних клітин, але в більшості випадків їх утворення супроводжується помітними морфологічними й структурними змінами: потовщується стінка вегетативної клітини, внаслідок чого утворюються оптично щільні структури, які сильно заломлюють світло (рис. 56).
У представників роду Azotobacter утворення цист супроводжується зміною морфології клітини, втратою джгутиків, накопиченням у цитоплазмі гранул ПОМК, синтезом додаткових покривів: зовнішніх (екзини) і внутрішніх (інтини). Ці оболонки (покриви) відрізняються між собою як структурно, так і за хімічнимскладом. Форми спокою деяких ціанобактерій називають акінетами. Вони більші за вегетативні клітини, мають подовжену або сферичну форму, товсту оболонку. Утворюються акінети в період затримки росту культури. Клітина при цьому збільшується в розмірах, у цитоплазмі накопичуються гранули запасних речовин (глікоген, поліфосфати, ціанофіцинові гранули), а також утворюються карбоксисоми. Одночасно потовщується пептидоглікановий шар, ущільнюється слизовий чохол. В цілому, оболонки акінети містять більше ліпідів і полісахаридів, а цитоплазма – менше води, ніж вегетативна клітина. В процесі формування акінет збільшується вміст ДНК, кількість рибосом, зменшується кількість хлорофілу і фікобілінових пігментів. Сформовані форми спокою протягом різного часу можуть зберігати свою життєздатність (табл. 10) і проростають у відповідних умовах з утворенням активних (у метаболічному відношенні) клітин.
Таблиця 10
|
|||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-07-16; просмотров: 785; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.142.130.127 (0.014 с.) |