Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Препарат 1. Семенник крысы (гистологический срез)Содержание книги
Поиск на нашем сайте
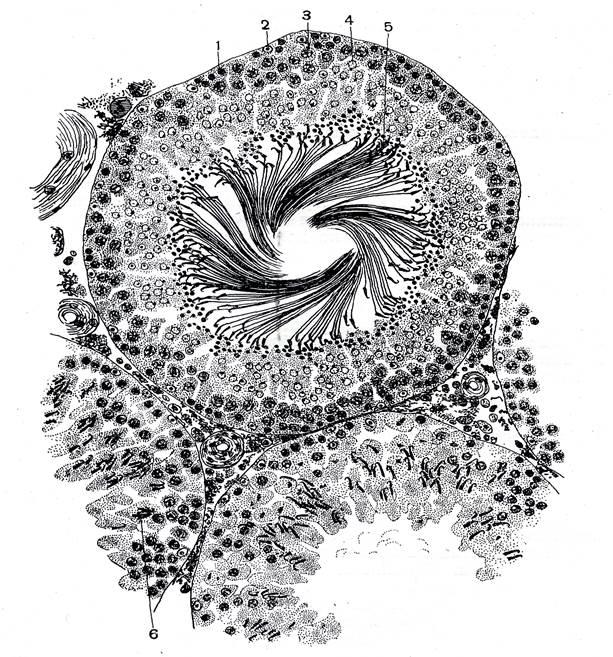
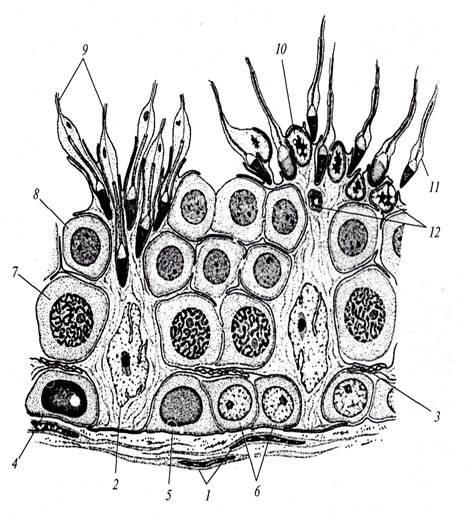
Препарат 1. Семенник крысы (гистологический срез) На срезе семенника канальцы выглядят либо округлыми, либо овальными – это зависит от плоскости сечения. Между семенными канальцами расположена соединительная ткань и клетки Лейдига, основная функция которых – выработка мужского полового гормона тестостерона, необходимого для осуществления сперматогенеза. Стенку канальца образуют несколько слоев клеток, которые представляют собой дифференцирующиеся половые клетки (рис. 1). Развитие их происходит в направлении от базальной мембраны в просвет канальца. Помимо сперматогенного эпителия (сперматогониев и их потомков) в состав стенки канальца входят соматические по происхождению клетки Сертоли. Основания этих клеток находятся на стенке канальца, а разветвленные их части простираются внутрь канальца. Рассмотреть границы клеток Сертоли невозможно, но в основании каждой клетки располагаются ядра треугольной формы. Функции клеток Сертоли различны: трофическая, опорная, регуляторная, фагоцитарная. Между основаниями этих клеток располагаются сперматогонии. Они характеризуются относительно небольшими размерами и крупным ядром. Хроматин в ядрах сперматогониев первой генерации деконденсирован, равномерно распределен в кариоплазме. Ярус семенного канальца (базальный компартмент), в котором расположены сперматогонии, обозначается зоной размножения. В околополостном отсеке ярусами располагаются последовательно сперматоциты первого порядка, спематоциты второго порядка и сперматиды. При формировании спермотозоидов от сперматид отделяются резидуальные тельца. В самом центре канальцев располагаются сперматозоиды. На рисунке 2 показана схема строения того же семенного канальца, позволяющая детально разобраться в строении сперматогенного эпителия.
Рис. 1. Поперечный разрез семенного канальца крысы (Мануилова, 1964). 1 – сперматогония; 2 – клетка Сертоли; 3 – сперматоцит первого порядка; 4 – сперматида; 5 – сперматозоид; 6 – преобразующаяся сперматида.
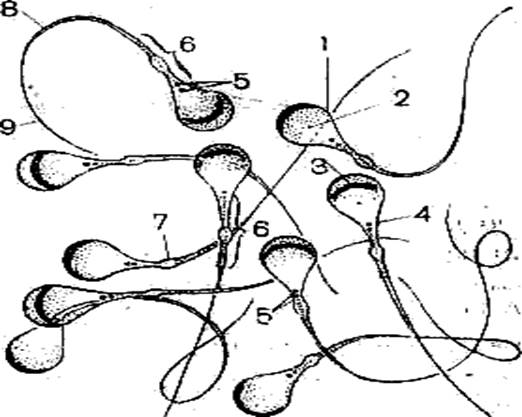
Рис. 2. Схема строения сперматогенного эпителия (Мануилова, 1964). 1 – ядра клеток оболочки семенного канальца; 2 – ядро клетки Сертоли; 3 – зоны плотных контактов между отростками соседних клеток Сертоли; 4,5,6 – сперматогонии; 7 – сперматоциты первого порядка; 8 – ранние (округлые) сперматиды; 9 – поздние (удлиненные) сперматиды; 10 – отделение резидуального тельца; 11 – сперматозоид в просвете семенного канальца; 12 – резидуальные тельца. Препарат 2. Сперматозоиды морской свинки Сперматозоиды морской свинки – это типичные жгутиковые клетки. Общая длина их равна 100 мкм. Отчетливо выражена головка, шейка и хвостик. Головка имеет округлую форму, центр ее занимает ядро. Акросома темная и в виде шапочки расположена на апикальном конце клетки. Область шейки несколько утолщена. Здесь сохраняется резидуальная цитоплазма в виде капельки. Капелька обычно исчезает по мере прохождения сперматозоида по канальцам придатка семенника. Хвостик длинный, покрыт цитоплазматической оболочкой, состоит их осевой нити и окружающей ее цитоплазмы. Иногда на препаратах можно увидеть «многохвостые» спермии. Это результат склеивания нескольких спермиев, которые обладают высокой способностью к агглютинации
.
Рис. 3. Сперматозоиды морской свинки (Новиков, Святенко, 1984). 1 – головка; 2 – ядро; 3 – акросома; 4 – шейка; 5 – две центриоли; 6 – связующий отдел; 7 – утолщение; 8 – главный отдел хвостика; 9 – концевой отдел хвостика.
Препарат 3. Сперматозоиды тритона Сперматозоиды тритона, как и морской свинки, относятся к жгутиковым формам. Размер клетки примерно равен 500 мкм. Головка сильно вытянута и заострена, в апикальной части имеет форму гарпуна. Под акросомой располагается крупное плотное стержневидное ядро. К задней его части примыкает очень короткая шейка. Цитоплазматическая оболочка длинного хвоста формирует ундулирующую мембрану, которая отсутствует лишь в самом дистальном конце.
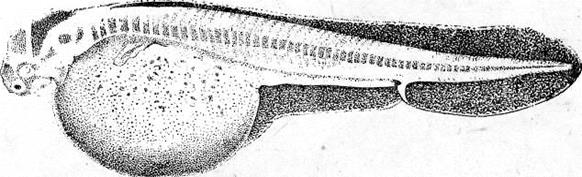
Рис. 4. Сперматозоид тритона (Газарян, Белоусов, 1983). Занятие 2 Оогенез Оогенезом называется образование женских половых клеток. Яйцеклетка – высокоспециализированная гаплоидная клетка, обладающая потенциальной способностью к развитию. В ее цитоплазме запасаются вещества, необходимые для роста и развития зародыша: структурные белки, ферменты, информационный материал, витамины, соли и др. В отличие от сперматозоида, яйцеклетки увеличивают объем цитоплазмы в десятки, а иногда и в тысячи раз. Основным питательным веществом, накапливаемым в цитоплазме, является желток. По количеству желтка яйцеклетки подразделяются на маложелтковые (олиголецитальные), со средним количеством желтка (мезолецитальные) и многожелтковые (полилецитальные). Иногда яйцеклетки почти лишены желтка (алецитальные). Распределение желтка в яйцеклетке также может быть различным. При равномерном распределении желтка, яйцеклетки называется изо- или гомолецитальными; если он смещен к одному из полюсов –телолецитальными. В случае если желток распределен в яйцеклетке неравномерно, то она приобретает полярность. Желток в большей степени откладывается на одном из полюсов (вегетативном), а на противоположном ему (анимальном) находится цитоплазма и сюда, как правило, смещается и ядро клетки. У членистоногих яйцеклетки полилецитальные, но желток расположен в центральной части цитоплазмы, ядро также расположено в центре (центролецитальные). Яйцеклетки, помимо плазматической оболочки, обычно имеют еще ряд оболочек. Так, сама яйцеклетка вырабатывает первичную или желточную оболочку. Вторичная оболочка формируется фолликулярными клетками, окружающими овулировавшее из яичника яйцо. Третичные оболочки, которые вырабатываются стенками яйцеводов, имеют наиболее сложное строение. Размеры и форма яиц варьируют в пределах животного царства. Как и при сперматогенезе, в оогенезе выделяются те же этапы, но отсутствует период формирования. В период размножения диплоидные оогонии совершают видоспецифичное число митотических деления, а затем переходят в стадию роста. Отличительной особенностью оогенеза является то, что размножающиеся ооциты обладают меньшей митотической активностью. У высших позвоночных пролиферация ооцитов осуществляется только в эмбриогенезе. В период роста в клетке осуществляются одновременно два процесса: в ядре – профаза первого мейотического деления, а в цитоплазме – накопление питательного и информационного материала. Рост ооцита делится на два периода: · период малого роста (превителлогенез); · период большого роста (вителлогенез). Накопление питательного материала в превителлогенезе происходит за счет собственного метаболизма клетки, а в вителлогенезе – за счет поступления его извне. В цитоплазме ооцитов желток запасается в виде гранул и представляет собой смесь компонентов (белки, жиры, углеводы), основным из которых является белок вителлогенин. У большинства позвоночных и некоторых беспозвоночных вителлогенин синтезируется в печени. В период вителлогенеза ооцит увеличивается в размерах в десятки, а иногда и в тысячи раз. Заканчивается оогенез делениями созревания. В результате первого деления происходит редукция числа хромосом, в результате второго – в дочерние клетки расходятся хроматиды. В результате формируются 4 гаплоидных клетки, но только одна из них становится зрелой яйцеклеткой. Итак, оогенез отличается от сперматогенеза рядом признаков: · число сперматогониальных делений всегда больше, чем оогониальных; · у высших позвоночных размножение оогониев происходит только в эмбриональный период; · период роста в оогенезе всегда более продолжительный, растущий ооцит увеличивается в размерах значительно, трофические компоненты клетки резервируются в цитоплазме; · в оогенезе отсутствует период формирования; · блок мейоза у разных животных происходит на разных фазах. Типы оогенеза Развитие женских половых клеток может осуществляться в любом участке тела (диффузный оогенез), либо в специализированных органах – яичниках (локализованный оогенез). Диффузный оогенез характерен для примитивных многоклеточных (губки), когда развивающиеся ооциты накапливают питательные вещества путем фагоцитоза близлежащих клеток. При локализованном оогенезе транспорт питательных веществ извне осуществляется специализированными клетками (алиментарный тип) или без участия последних (солитарный тип). При солитарном накоплении питательных веществ все необходимые макромолекулы ооцит получает из полости гонады в виде простых низкомолекулярных соединений. Алиментарный тип оогенеза может быть фолликулярным или нутриментарным. При фолликулярном оогенезе растущий ооцит окружен фолликулярными клетками (соматическими по происхождению), которые вместе с ним образуют функциональную структуру – фолликул. Это наиболее распространенный тип оогенеза, происходящий у всех хордовых. При нутриментарном оогенезе ооцит окружен трофоцитами (клетками-кормилками), связанными с ним цитоплазматическими мостиками. Трофоциты – абортивные половые клетки, имеющие общее происхождение с развивающимся ооцитом. Нутриментарный оогенез описан для высших червей и насекомых. Яичник млекопитающих – это плотный орган, строма которого представлена соединительной тканью. На поперечных срезах различимо корковое вещество (cortex), в котором находятся фолликулы всех стадий развития, и мозговое вещество (medulla), где проходят крупные сосуды яичника. Стенка яйцевых фолликулов образована соединительнотканной текой, в которой разветвляются капилляры, питающие фолликул, и фолликулярным эпителием. Впервые половая клетка устанавливает связь с фолликулярными, когда вступает в период малого роста и превращается в ооцит 1 порядка В это время в ее ядре начинается профаза мейоза, но не завершается, а блокируется на стадии диплотены. Так образуются примордиальные или первичные фолликулы (рис. 6).. Они представляют собой резерв половых клеток, которые возобновляют свое развитие при половом созревании. Располагаются они, как правило, группами в самых поверхностных участках корковой зоны яичника и являются генерацией покоящихся фолликулов. Примордиальные фолликулы порциями вступают в рост в течение всей жизни самки. При инициации развития фолликулярные клетки начинают пролиферировать, из плоских превращаются в кубические, а затем и призматические, и начинают покрывать ооцит первого порядка сначала одним слоем, а затем двумя, тремя и др. Фолликулярная оболочка многослойного фолликула называется зернистой (zona granulosa), снаружи от нее начинает формироваться соединительнотканная оболочка (teca folliculi). После того как вокруг ооцита первого порядка завершается формирование одного слоя фолликулярных клеток, начинается формирование блестящей оболочки ооцита (zona pellucida). Эту оболочку пронизывают микровыросты ооцита и макровыросты фолликулярных клеток. Первичный фолликул увеличивается в размерах, как за счет собственного роста, так и за счет увеличения количества слоев фолликулярных клеток. 4 6 Рис. 7. Граафов пузырек из яичника кошки (Мануилова, 1964). 1 – яйцо; 2 – яйценосный бугорок; 3 – блестящая оболочка; 4 – тека; 5 – полость пузырька; 6 – фолликулярный эпителий. Ооцит окружен блестящей оболочкой (zona pellucida) и лучистым венцом (corona radiata). Полость фолликула ограничена зернистой оболочкой (zona granulosa). Снаружи фолликул покрыт базальной мембраной и текальной оболочкой, состоящей из внутренней части, содержащей сосуды, и наружной, в состав которой входят стромальные и мышечные клетки. Граафов пузырек на завершающих стадиях своего развития быстро увеличивается в размерах, приближается к поверхности яичника, и его верхушка (стигма) выдается над ней. Впоследствии происходит разрыв cтенки в области стигмы и овуляция ооцита. На месте овулировавшего ооцита, оставшиеся фолликулярные клетки претерпевают железистый метаморфоз и превращаются в желтое тело. Занятие 3 Оплодотворение В зрелых половых клетках практически все жизненные процессы подавлены. Клетки фактически находятся в состоянии анабиоза, и только оплодотворение у большинства видов животных является толчком к дальнейшему развитию. Оплодотворение представляет собой процесс слияния генетического материала двух половых клеток (яйцеклетки и сперматозоида), в результате чего возникает новая особь с потенциями обоих родителей. Таким образом, оплодотворение осуществляет две основные функции: – половую, когда комбинируются гены двух родителей; – репродуктивную, когда создается новый организм. Всякому оплодотворению предшествует процесс осеменения. Осеменение может быть наружным и внутренним. Наружное осеменение свойственно большинству водных животных или тем, кто размножается в воде. У животных, размножающихся на суше, как правило, оплодотворение внутреннее. При осеменении у животных и человека происходит выделение большого количества сперматозоидов. Среда женских половых путей не очень благоприятна для сперматозоидов, и миллионы их погибают, не достигнув яйцеклетки. Дистантное взаимодействие гамет обеспечивает вероятность встречи сперматозоида с яйцеклеткой. При наружном оплодотворении – это видоспецифическое привлечение спермиев своего вида и их активация посредством хемотаксисов. При внутреннем оплодотворении, происходящем (например, у человека), в верхней трети яйцеводов, основная роль в дистантном взаимодействии гамет отводится явлению реотаксиса – способности спермиев двигаться против встречного течения жидкости в маточных трубах. Контактное взаимодействие гамет складывается из нескольких реакций. Первая из них – акросомная. Сначала происходит слияние наружной мембраны акросомы с плазматической мембраной сперматозоида, а затем – разрыв этих мембран, сопровождающийся экзоцитозом содержимого акросомного пузырька. Сперматолизины, являющиеся основным компонентом акросомы, растворяют третичную оболочку яйцеклетки. Благодаря выходу ионов водорода из головки спермия происходит образование акросомного выроста. Акросомный вырост вступает в контакт с наружной поверхностью яйцеклетки – желточной оболочкой. Белок биндин отвечает за специфическую реакцию взаимодействия мембран яйцеклетки и спермия и встроен в головку спермия. Далее происходит активация метаболизма яйцеклетки. В месте проникновения спермия на поверхности яйцеклетки образуется бугорок оплодотворения. Формирование его сопровождается активацией белка актина яйцеклетки. Мембраны акросомной нити и яйцеклетки сливаются, и через образовавшийся цитоплазматический мостик происходит слияние цитоплазмы спермия и ооцита (плазмогамия), а также проникновение сперматического ядра в ооплазму. Участок мембраны сперматозоида встраивается в мембрану яйцеклетки. После контакта сперматозоида с плазматической мембраной яйцеклетки в первые секунды происходит блок полиспермии (у большинства групп животных). В яйцеклетке происходит высвобождение ионов кальция из клеточных депо, а это в свою очередь приводит к кортикальной реакции и образованию оболочки оплодотворения. Полное завершение мейоза без участия сперматозоида, происходит лишь у немногих видов животных (некоторые кишечнополостные, некоторые иглокожие). У большинства видов мейоз не завершается и блокируется в конце профазы (как правило, на стадии диплотены-диакинеза). Это – первый блок мейоза, который у позвоночных может продолжаться годами и снимается во время овуляции. Однако и после этого часто мейоз не доходит до конца – наступает второй блок мейоза, краткосрочный. Для завершения мейоза необходимо снять этот блок активирующим агентом. В роли такого агента выступает сперматозоид. Всех животных можно поделить на группы в зависимости от того, как эти остановки мейоза накладываются на процессы овуляции и оплодотворения: · мейоз останавливается один раз – на стадии диакинеза и снимается при оплодотворении (губки, некоторые представители плоских, круглых и кольчатых червей, моллюски); · первый блок мейоза наступает на стадии диплотены-диакинеза профазы мейоза и снимается во время овуляции; второй блок мейоза наступает на метафазе первого деления созревания и снимается оплодотворением (некоторые губки, немертины, моллюски, практически все насекомые; сюда можно отнести и некоторых млекопитающих – собаки, лисицы, лошади); · первый блок мейоза наступает на стадии диплотены-диакинеза профазы мейоза в эмбриональный период развития и снимается овуляцией; второй блок мейоза наступает на стадии метафазы второго деления созревания, т. е. уже после выделения первого полярного тельца (практически все хордовые). У большинства животных сперматозоид входит в яйцеклетку целиком, включая и хвостик. Но даже если жгутик и окружающие его митохондрии проникли в яйцеклетку, они не играют никакой роли при дальнейшем развитии. Попав в яйцеклетку, сперматическое ядро поворачивается на 180°, и дистальная центриоль встает впереди него, осуществляя передвижение ядра в ооплазме. Затем сперматическое ядро преобразуется в мужской пронуклеус. В это время в яйцеклетке завершаются деления созревания и пронуклеусы совершают сложные движения (танец пронуклеусов) навстречу друг другу. Занятие 4 Дробление и бластуляция Дробление – самый ранний период развития зародыша, во время которого оплодотворенное яйцо делится на большое количество клеток. В период дробления митотический цикл имеет малую продолжительность, в нем не наблюдаются пре- и постсинтетические фазы, отсутствует постмитотический рост клеток. Умножение числа мелких клеток приводит к огромному увеличению их общей площади. В ходе митотического дробления в геометрической прогрессии увеличивается содержание и синтез ДНК, восстанавливается ядерно-цитоплазматическое соотношение, которое нарушается в период вителлогенеза ооцита. Дочерние клетки, образующиеся в период дробления, называются бластомерами, а делящие их плоскости – бороздами дробления. Борозды, проходящие через анимальный и вегетативный полюса зиготы, называются меридиональными. Борозды, проходящие перпендикулярно к полярной оси зиготы, проходящей через ее середину, называются экваториальными. Борозды, смещенные к анимальному полюсу, получили название широтных; а борозды, проходящие параллельно поверхности зародыша, называются тангенциальными. На начальных этапах дробления происходит синхронное деление всех бластомеров. Впоследствии появляется и нарастает асинхронность. На расположение ядер, веретен дробления и бластомеров влияют количество и расположение желтка в цитоплазме яйцеклеток. Желток, будучи необходимым питательным материалом для развития зародыша, является и пассивным балластом, задерживающим деление зиготы и бластомеров. Чем больше желточных гранул, тем медленнее делится эта часть цитоплазмы. Правила зависимости между расположением желтка и положением ядер и веретен были сформулированы немецким ученым О. Гертвигом на основе правил, предложенных ранее ботаником Ю. Саксом для растительных меристем. Суть этих правил состоит в следующем: · ядро стремится расположиться в центре свободной от желтка цитоплазмы; · веретено деления располагается перпендикулярно к наибольшему протяжению свободной от желтка цитоплазмы. На таблице 1 показана связь типов дробления яиц с количеством и распределением в них желтка. Каждый тип дробления следует рассматривать на конкретном примере: полное, более или менее равномерное встречается у морского ежа; полное неравномерное – у амфибий и хрящевых ганоидов; неполное дискоидальное – у костистых рыб, рептилий и птиц. Яйцеклетки с небольшим и средним количеством желтка (олиго- и мезолецитальные) претерпевают полное (голобластическое) дробление. При этом содержимое зиготы входит в состав бластомеров и зародыш развивается за счет внутриклеточной утилизации желтка. Полное дробление может быть равномерным (олиголецитальные яйца) и неравномерным (мезолецитальные яйца). Яйцеклетки с большим количеством желтка (полилецитальные) претерпевают неполное (меробластическое) дробление. Меробластический тип дробления характеризуется тем, что зародыш развивается лишь из небольшой части яйцеклетки, а остальная часть ее – питательный материал. Неполное дробление может быть дискоидальным (телолецитальные яйцеклетки с большим количеством желтка) и поверхностным (центролецитальные яйцеклетки). Характер дробления зависит не только от количества и расположения желтка, но и от ряда других факторов, в частности, от расположения кортикальных гранул в цитоплазме яйцеклетки. Так, дробление может быть радиальным, спиральным, билатеральным, анархичным и т.п. При радиальном дроблении плоскости последовательных дроблений проходят перпендикулярно друг к другу, и бластомеры располагаются симметрично относительно любой плоскости, проходящей через анимально-вегетативную ось яйца. При спиральном дроблении происходит смещение (поворот) бластомеров относительно оси яйца. Это вызывает скручивание веретена деления в анафазе. Плоскости веретен образуют спирали или участки спиралей, и бластомеры располагаются не правильными рядами, а более или менее чередуются друг с другом. Заключительный этап дробления – бластуляция характеризуется появлением ряда новых морфологических особенностей зародыша – бластодермы, первичной полости тела (бластоцели); новых биохимических функций в зародышевых клетках: начало работы зародышевого генома, начало синтеза РНК, белка. Бластоцель – это первый отсек внутренней среды организма, отличается по своему ионному составу от окружающей среды. Изоляция бластоцели от наружной среды обеспечивается формированием на боковых поверхностях соседних бластомеров специализированных плотных межклеточных контактов, непроницаемых для ионов. Проникновение ионов в бластоцель и обратно осуществляется через клетки бластодермы. Занятие 5 Занятие 6 Развитие хрящевых ганоидов (осетровых рыб) Хрящевым ганоидам свойственен тот же примитивный тип развития, какой характерен для амфибий, а также круглоротых, костных ганоидов, кистеперых и двоякодышащих рыб. У осетровых рыб умеренно телолецитальные яйца, полное неравномерное дробление, непрямое развитие с личиночной стадией и метаморфозом. Однако, яйца хрящевых ганоидов более дифференцированы в отношении разделения желточного запаса и образовательной цитоплазмы. Неравномерность в дроблении проявляется в том, что некоторые борозды лишь врезаются в яйцо, не достигая вегетативного полюса, а другие делят яйцо полностью. В результате формируется амфибластула. Мелкие анимальные бластомеры занимают сначала 1/3 поверхности яйца. Гаструляция похожа на таковую амфибий и проявляется в инвагинации и эпиболии. При инвагинации мелкие клетки анимального полушария проникают внутрь через дорзальную и латеральные губы бластопора и формируют крышу архентерона. Здесь располагаются зачатки хорды и мезодермы. Клетки, проходящие внутрь через вентральную губу бластопора, образуют дно архентерона с энтодермальной подушкой. Эпиболия проявляется в наползании микромеров на вегетативные макромеры. Когда микромеры покрывают еще только половину поверхности яйца, инвагинация уже заканчивается, и эти микромеры, лежащие у дорзальной губы бластопора, являются материалом для образования нервной пластинки. Вслед за окончанием гаструляции этот материал образует два сближенных между собой медуллярных валика. К концу обрастания желточной пробки, передний конец медуллярных валиков раздвигается, образуя передний и задний мозговые пузыри. В то же время нервные валики в задней части удлиняются и задним краем нарастают на желточную пробку. К этому моменту начинается сегментация сомитов, в передних из них начинают формироваться пронефросы. После закрытия желточной пробки формирование туловищных сомитов завершается, а вольфовы протоки достигают полной длины. В головной части формируются мозговые пузыри, которые значительно выступают над поверхностью яйца, две пары жаберных щелей (их число затем возрастает до пяти пар), слуховые пузырьки. На заднем конце зародыша в виде бугорка появляется хвостовая почка. Все эти образования распластаны на сферической поверхности яйца. Затем, по мере роста и дифференцировки сомитов, головной отдел зародыша и передние 20 сегментов туловища приподнимаются над поверхностью яйца (желтка). Хвостовая почка увеличивается в длину. На поверхности желтка под передним концом головы образуется сердце. По мере потребления желтка энтодермальной подушки его размеры уменьшаются только в дорзовентральном направлении. Снизу головного отдела и в области 40-го сомита, путем образования наружных выемок и полостей возникают ротовое и анальное отверстия. Происхождение последнего никак не связано с бластопором, который замкнулся на уровне 20-го сомита. В образовании хвостовой почки принимают участие продолжения зачатков всех осевых органов, собирающихся в области бластопора, превратившегося при замыкании нервной трубки в нервно-кишечный канал. При продолжении роста хвостовой почки и достижения зародышем предельной длины, свойственной данному виду, рост зародыша в длину прекращается и нервно-кишечный канал зарастает. Из яйца осетровых рыб развивается личинка, которая имеет внутренние жабры, торчащие из-под жаберной крышки, короткую голову, обонятельные ямки и недоразвитые глаза. По краям рта возникают мелкие острые зубы, сходные по строению с зубами акул. Есть зачатки наружных жабр и брызгальца. Тело личинки сверху и снизу до конца желточного мешка окаймлено плавательной эктодермальной складкой. Личинка имеет зачатки грудных плавников в виде небольших полукруглых складок. Метаморфоз протекает быстро. Образуются жаберные крышки, вырастает рострум. Размеры желточного мешка значительно сокращаются. Формируются спинные, грудные, брюшные и хвостовой плавники. Тело малька покрывается чешуёй (жучками) и появляются накладные кости черепа. Как и у амфибий, метаморфоз у хрящевых ганоидов связан с активностью щитовидной железы.
А. Б.
В. Г. Д. Е.
Ж.
З.
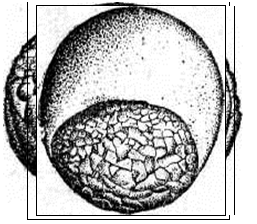
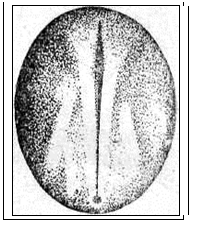
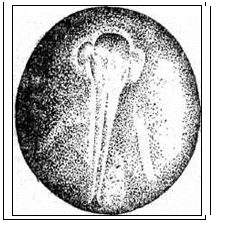
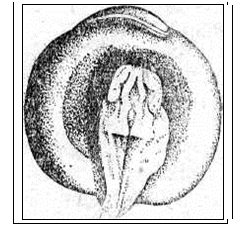
Рис. 20. Развитие хрящевых ганоидов (Иванов, 1945). А - начало дробления; Б - бластула; В - начало гаструляции; Г - конец гаструляции; Д - нейрула; Е - обрастание желтка зародышем; Ж - отделение зародыша от желтка; З - личинка. Занятие 7 Развитие птиц Птицы являются представителями высших позвоночных, эмбрионез которых приспособлен к наземным условиям. С развитием птиц лучше всего знакомиться на примере домашней курицы – классическом объекте изучения закономерностей эмбрионального развития. Яйцеклетки птиц – полилецитальные. Зародышевая плазма занимает небольшой участок на анимальном полюсе и представлена зародышевым диском, резко отграниченным от желтка. Собственно яйцеклеткой является то, что в общежитии называют “желтком”. В виде голого желтка она выходит из яичника. Сразу же по выходе, в самых начальных частях яйцевода происходит полиспермное оплодотворение с образованием желточной оболочки. Проходя по яйцеводу к клоаке, оплодотворенное яйцо окружается в различных отделах яйцевода белком. В конечном отделе яйцевода образуется скорлупа. Продолжительность развития куриного эмбриона – 21 день. Дробление в курином яйце начинается сразу после оплодотворения, протекает в яйцеводе и продолжается по мере того, как яйцеклетка одевается белковой оболочкой и скорлупой. После откладки яйца до насиживания дальнейшего развития не происходит. Возобновляются процессы развития с момента насиживания. Дробление меробластическое, дискоидальное. Дискобластула представляет собой многоклеточную пластинку, распластанную на желтке, причем между зародышевым диском и желтком обычно имеется небольшая полость – бластоцель. Гаструляция состоит из двух фаз и происходит еще в то время, когда яйцо находится в половых путях самки. I фаза - расслоение (деляминация) зародышевого диска на эпибласт и гипобласт. Впоследствии из первого образуются зародышевые листки и большая часть внезародышевого материала, а из второго – часть внезародышевой энтодермы. Кроме того, в начале инкубации в центре зародышевого диска (эпибласта) выделяется зародышевый щиток. Окружающая его часть зародышевого диска оказывается к этому времени несколько приподнятой над желтком. Эта часть носит название светлого поля - area pellucida (клетки будущего зародыша). Светлое поле окружено плотно прилежащей к желтку бластодермой, получившей название темного поля - area opaca (клетки будущих внезародышевых, провизорных органов: амниона, серозы, аллантоиса). Наружный край темного поля (гипобласта) является краем обрастания. II фаза гаструляции заключается в активных клеточных перемещениях в эпибласте, приводящих к образованию всех трех зародышевых листков как в зародышевой, так и во внезародышевых частях, которые происходят в первые сутки инкубации. Главнейшие перемещения совершаются в области зародышевого щитка. Перемещение клеточного материала здесь идет в направлении от передного к заднему концу будущего зародыша. Клеточные потоки особенно быстро движутся по краям зародышевого щитка и быстрее достигают заднего его края. Два клеточных потока, встречаясь по срединной линии, направляются вперед. Клеточный материал медиальной части движется медленнее. Боковые клеточные потоки в срединной части образуют утолщенную клеточную полоску, которая постепенно удлиняется от задней части зародышевого щитка к передней. Это образование получило название первичной полоски. В месте встречи клеточных потоков, движущихся с переднего и заднего концов зародышевого щитка, образуется бугорок, возвышающийся над зародышевым щитком - первичный или гензеновский узелок. На вершине его появляется углубление - первичная ямка, а по середине первичной полоски возникает первичная бороздка, являющаяся продолжением первичной ямки. Передний край первичной ямки является дорзальной губой бластопора, а края первичной бороздки - латеральными губами. Затем передний край первичной ямки надвигается на лежащий кзади клеточный материал так, что бластопор начинает замыкаться спереди назад и находящийся непосредственно перед первичным узелком клеточный материал подворачивается через дорзальную губу бластопора, формируя между наружным и внутренним листками хордальный или головной отросток. Одновременно, материал первичной полоски начинает погружаться внутрь через латеральные губы бластопора. Он проникает между наружным и внутренним зародышевыми листками, ложась по бокам хордального отростка, распространяясь в стороны и вперед. После гаструляции снаружи остаются только материал нервной пластинки и кожная мезодерма. Участки мезодермальных выростов, расположенные в передних частях первичной полоски, лежащие по сторонам от хордального выроста, дают сомиты. Мезодермальный материал, лежащий в задних частях первичной полоски, смещается вперед позже и входит в состав спланхнотомов. Таким образом, возникают два мезодермальных крыла, свободные края которых разрастаются за пределы зародышевого щитка и вклиниваются между экто- и энтодермой зародышевого диска, продолжаясь во внезародышевую часть. Материал, лежащий снаружи впереди и по бокам от гензеновского узелка, параллельно с образованием хордального отростка, также вытягивается по осевой линии в длину и дает начало нервной пластинке. Формирование зачатков органов и отделение клеточного материала сходно с таковым у ранее изученных групп позвоночных животных. Энтодерма первоначально представлена плоской пластинкой клеток, распластанной над заполненной жидкостью подзародышевой полостью. Лишь позднее зародыш начинает приподниматься над желтком, энтодермальная пластинка выгибается кверху, образуя под хордой кишечный желобок, который позже сворачивается в кишечную трубку. Образование ее начинается на переднем, а затем на заднем конце тела и постепенно распространяется от обоих концов к середине. Как и у хрящевых ганоидов, у птиц формируется желточный мешок, выполняющий функцию переваривания и всасывания желтка, и служащий на ранних этапах развития кроветворным органом и органом дыхания. Нервная пластинка образует желобок, а затем свертывается в трубку, погружаясь под кожную эктодерму. Мезодерма дифференцируется на сомиты, занимающие наиболее осевое положение по бокам хорды и нервной трубки, а также, лежащие латеральнее - нефротомы. Наиболее латерально расположенные участки мезодермы - спланхнотомы. Сомиты и нефротомы представляют сегментированную часть мезодермы, причем сегментация распространяется спереди назад. На более поздних стадиях развития сомиты подразделяются на дерматомы, миотомы и склеротомы, из которых развиваются, соответственно, кориум кожи, мышцы и дефинитивный осевой скелет в виде материала позвоночника, расположенного вокруг хорды. Одновременно, спланхнотомы, расщепившись на париетальный (наружный) и висцеральный (внутренний) листки, образуют целом. Нефротомы дают начало вольфову протоку и канальцам первичной почки. Тело зародыша отделяется от внезародышевой части туловищной складкой. Внезародышевая часть разделена на две пластинки экзоцеломом. Наружная пластинка состоит из эктодермы и париетальной мезодермы и поднимается над телом зародыша в виде амниотической складки. Внутренняя пластинка состоит из висцеральной мезодермы и энтодермы, образуя стенку желточного мешка. В мезодерме видны кровеносные сосуды. Список рекомендуемой литературы 1. Алмазов И. В., Сутулов Л.С. Атлас по гистологии и эмбриологии. М., 1978, 544 с. 2. Белоусов Л. В. Введение в общую эмбриологию. М., 1980. 211 с. 3. Газарян К. Г., Белоусов Л. В. Биология индивидуального развития животных. М., 1983. 287 с. 4. Голиченков В.А., Иванов Е.А., Лучинская Н.Н. Практикум по эмбриологии. М., 2004. 205 с. 5. Иванов П. П. Руководство по общей и сравнительной эмбриологии. Л., 1945. 6. Кнорре А. Г. Краткий очерк эмбриологии человека. Л.,1967. 7. Мануилова Н.А. Гистология с основами эмбриологии. М., 1964. 8. Новиков А. И., Святенко Е. С. Руководство к лабораторным занятиям по гистологии с основами эмбриологии. М.,1984. 9. Токин Б. П. Общая эмбриология. М., 1987. 480 с. 10. Фалин Л. И. Атлас микрофотографий по нормальной гистологии и эмбриологии. М., 1957.
«Биология размножения и развития». Учебно-методическое пособие для студентов очной и очно-заочной формы обучения по специальности «020201 – Биология»
Составители: Галина Анатольевна Фадеева Ульяна Вениаминовна Колова Светлана Анатольевна Мельник
|
||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-06-22; просмотров: 3855; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.26.184 (0.019 с.) |