Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Цикл кальвина можно разделить на фазы.Содержание книги Поиск на нашем сайте
Первая фаза — карбоксилирование. Эта реакция катализируется специфическим для процесса фотосинтеза ферментом рибулозобисфосфат-карбоксилазой/ оксигеназой (сокращенно РБФ-карбоксилаза/оксигеназа), в научной литературе последних лет чаще встречается под названием Rubisco, от ribulosobiphosphatecarboxylase/oxygenase. Впервые Rubisco был выделен и очищен в 1955 году. Особенностью фермента является то, что катализируемая им реакция является самой медленной стадией в цикле фиксации углекислоты. В листьях Rubisco содержится в больших количествах и является основной фракцией белка хлоропластов. Более того, высказывается мнение, что это самый распространенный белок-фермент на земном шаре. В активное состояние фермент переходит при освещении хлоропластов. Уже отмечалось, что формирование этого фермента происходит под контролем двух геномов: большие субъединицы (54 кДа) кодируются в ядре, синтезируются в цитоплазме; малые — кодируются и синтезируются в хлоропласте. При взаимодействии РБФ с С02образуется сначала промежуточное нестойкое шестиуглеродное соединение, которое затем распадается на две молекулы ФГК. Образовавшаяся ФГК — это органическая кислота, и ее энергетический уровень ниже уровня Сахаров. Поэтому это соединение не может непосредственно превращаться в углеводы. Необходимо превращение его в трехуглеродный сахар — фосфоглицериновый альдегид (ФГА). Вторая фаза — восстановление. Дальнейшие превращения ФГК требуют участия продуктов световой фазы фотосинтеза: АТФ и НАДФН + Н+. Реакция идет в два этапа. Прежде всего, происходит реакция фосфорилирования 3-ФГК. Донором фосфатной группы является АТФ. АТФ требуется здесь в качестве дополнительного источника энергии. Образуется 1,3-дифосфоглицериновая кислота. Реакция катализируется ферментом фосфоглицерокиназой:
Образовавшееся в этой реакции соединение — дифосфоглицериновая кислота — обладает более высокой реакционной способностью, содержит макроэргическую связь, полученную от АТФ. Затем карбоксильная группировка этого соединения восстанавливается до альдегидной с помощью триозофосфатдегидрогеназы, коферментом которой служит НАДФ. Образовавшийся ФГА является по уровню восстановленным углеводом. Это соединение вступает в две последние фазы. Пять молекул ФГА используется на регенерацию акцептора РБФ для того, чтобы фиксация С02могла снова осуществляться. Оставшаяся шестая молекула вступает в фазу «синтеза продуктов», где превращается в более сложные соединения (углеводы, аминокислоты и др.). Третья фаза — регенерация. В процессе регенерации акцептора используется пять молекул ФГА, в результате чего образуются три молекулы рибулезо-5-фос-фата. Этот процесс идет через образование 4-, 5-, 6-, 7-углеродных соединений. Прежде всего, первая молекула ФГА изомеризуется до фосфодиоксиацетона. Процесс катализируется ферментом триозофосфатизомеразой. От ФДФ отщепляется фосфат, и ФДФ превращается во фруктозо-6-фосфат (ф-6-Ф). Далее от Ф-6-Ф (С6) отщепляется 2-углеродный фрагмент (—СО—СН2ОН), который переносится на следующую (третью) триозу. Это транскетолазная реакция идет при участии фермента транскетолазы. В результате образуется первая пентоза (С5)-рибулезофосфат. От Ф-6-Ф остается 4-углеродный сахар эритрозофосфат фосфорилирование. Для этого используются три молекулы АТФ, образовавшиеся в результате световых реакций. Все реакции, входящие в цикл Образовавшийся триозофосфат (ФГА) вступает в четвертую стадию темновых реакций — стадию образования продуктов фотосинтеза. (С4), который конденсируется с четвертой триозой с образованием седогептулезодифосфата (С7). После отщепления фосфата седогептулезодифосфат превращается в седогептулезофосфат. Далее снова происходит транскетолазная реакция, в результате которой от седогептулезофосфата отщепляется 2-углеродный фрагмент, который переносится на пятую триозу Образуются еще две молекулы рибулезофосфата. Таким образом, в результате рассмотренных реакций получаются 3 молекулы рибулезофосфата. При прохождении двух циклов из 12 молекул образовавшегося ФГА две молекулы выходят из них, образуя одну молекулу фруктозодифосфата (ФДФ). Из двух молекул фруктозодифосфата (Ф-1, 6-диФ) образуются фруктозо-6-фосфат (ф-6-Ф) и глюкозо-1-фосфат (Г-1-Ф). Глюкозо-1-фосфат, взаимодействуя с уридинтрифосфатом (УТФ), дает уридиндифосфоглюкозу (УДФГ). В свою очередь УДФГ, реагируя с Ф-6-Ф, дает сахарозофосфат. По-видимому, именно сахароза является первым свободным сахаром, образующимся в процессе фотосинтеза. Из сахарозы образуются нефосфорилированные моносахара (глюкоза и фруктоза). Крахмал образуется из аденозиндифосфоглюкозы (АДФГ) или уридиндифосфоглюкозы (УДФГ), процесс катализируется ферментом амилосинтетазой. Среди первых продуктов фотосинтеза обнаружены аминокислоты. По-видимому, ФГК, образовавшаяся на первом этапе цикла Кальвина, может превращаться в пировиноградную кислоту. Этот процесс идет особенно интенсивно при недостатке НАДФН, из-за чего задерживается преобразование ФГК в ФГА (обычный путь в цикле Кальвина). Пировиноградная кислота в присутствии NH3 дает аминокислоту аланин. Показано, что скорость включения 14С02 в аланин в клетках хлореллы при некоторых условиях может даже превышать скорость ее включения в сахарозу. Из пировиноградной кислоты может образоваться еще ряд органических кислот (в цикле Кребса).
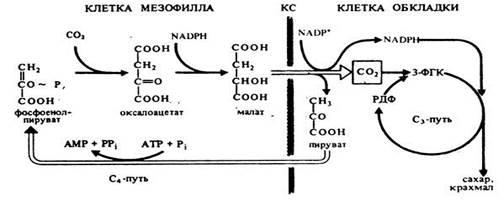
С4 путь фотосинтеза
С4-путь фотосинтеза или цикл Хетча-Слэка Австралийскими учеными М. Хетчем и К. Слэком был описан С4-путь фотосинтеза, характерный для тропических и субтропических растений однодольных и двудольных 16 семейств (сахарный тростник, кукуруза и др.). Большинство самых злостных сорняков – С4 растения, а большинство сельскохозяйственных культур относятся к С3-растениям. Листья этих растений содержат хлоропласты двух типов: обычные в клетках мезофилла и крупные хлоропласты, не имеющие гран и фотосистемы II, в клетках обкладки, окружающих проводящие пучки. В цитоплазме клеток мезофилла фосфоэнолпируваткарбоксилаза присоединяет СО2 к фосфоэнолпировиноградной кислоте, образуя щавелевоуксусную кислоту. Она транспортируется в хлоропласты, где восстанавливается до яблочной кислоты при участии НАДФН (фермент НАДФ+-зависимая малатдегидрогеназа). В присутствии ионов аммония щавелевоуксусная кислота превращается в аспарагиновую кислоту (фермент - аспартатаминотрансфераза). Яблочная и (или) аспарагиновая кислоты переходят в хлоропласты клеток обкладки, декарбоксилируются до пировиноградной кислоты и СО2. СО2 включается в цикл Кальвина, а пировиноградная кислота переносится в клетки мезофилла, где превращается в фосфоэнолпировиноградную кислоту. В зависимости от того, какая кислота – малат или аспартат – транспортируется в клетки обкладки, растения делят на два типа: малатный и аспартатный. В клетках обкладки эти С4-кислоты декарбоксилируются, что происходит у разных растений происходит с участием различных ферментов: НАДФ+-зависимой малатдегидрогеназы декарбоксилирующей (НАДФ+-МДГ), НАД+-зависимой малатдегидрогеназы декарбоксилирующей (малик-энзим, НАД+-МДГ) и ФЭП-карбоксикиназы (ФЕП-КК). Поэтому растения делят еще на три подтипа: НАДФ+-МДГ-растения, НАД+-МДГ-растения ФЕП-КК-растения. Такой механизм позволяет растениям фотосинтезировать при закрытых из-за высокой температуры устьицах. Кроме того, продукты цикла Кальвина образуются в хлоропластах клеток обкладки, окружающих проводящие пучки. Это способствует быстрому оттоку фотоассимилятов и тем самым повышает интенсивность фотосинтеза. Фотосинтез по типу толстянковых (суккулентов)-САМ путь. В сухих местах существуют растения-суккуленты, у которых устьица открыты ночью и закрыты днем для уменьшения транспирации. В настоящее время этот тип фотосинтеза обнаружен у представителей 25 семейств. У суккулентов (кактусов и растений сем. толстянковых (Crassulaceae) процессы фотосинтеза разделены не в пространстве, как у других С4-растений, а во времени. Этот тип фотосинтеза получил название CAM (crassulation acid metabolism)-путь. Устьица днем обычно закрыты, что предотвращает потерю воды в ходе транспирации, и открыты ночью. В темноте СО2 поступает в листья, где фосфоэнолпируваткарбоксилаза присоединяет его к фосфоэнолпировиноградной кислоте, образуя щавелевоуксусную кислоту. Она восстанавливается НАДФН-зависимой малатдегидрогеназой до яблочной кислоты, которая накапливается в вакуолях. Днем яблочная кислота переходит из вакуоли в цитоплазму, где декарбоксилируется с образованием СО2 и пировиноградной кислоты. СО2 диффундирует в хлоропласты и включается в цикл Кальвина. Итак, темновая фаза фотосинтеза разделена во времени: СО2 поглощение ночью, а восстанавливается днем, из ЩУК образуется малат, карбоксилирование в тканях происходит дважды: ночью карбоксилируется ФЕП, днем РуБФ. В САМ-растения делят на два типа: НАДФ-МДГ-растения, ФЕП-КК-растения. Как С4, САМ-тип является дополнительным, поставляющим СО2 в С3-цикл у растений, приспособившихся к жизни в условиях повышенных температур или недостатка влаги. У некоторых растений этот цикл функционирует всегда, у других – только в неблагоприятных условиях. Фотодыхание. Фотодыхание – это активируемый светом процесс выделения СО2 и поглощения О2.(НИ К ФОТОСИНТЕЗУ,НИ К ДЫХАНИЮ НЕ ОТНОСИТСЯ). Так как первичным продуктом фотодыхания является гликолевая кислота, оно еще называется гликолатным путем. Фотодыхание усиливается при низком содержании СО2 и высокой концентрации О2 в воздухе. В этих условиях рибулозобисфаткарбоксилаза хлоропластов катализирует не карбоксилирование рибулозо-1,5-дифосфата, а его расщепление на 3-фосфоглицериновую и 2-фосфогликолевую кислоты. Последняя дефосфорилируется с образованием гликолевой кислоты. Гликолевая кислота из хлоропласта переходит в пероксисому, где окисляется гликолатоксидазой до глиоксиловой кислоты. Образующаяся при этом перекись водорода разлагается каталазой, присутствующей в пероксисоме. Глиоксиловая кислота аминируется, превращаясь в глицин. Глицин транспортируется в митохондрию, где из двух молекул глицина синтезируется серин и освобождается СО2. Серин может поступать в пероксисому и под действием аминотрансферазы передает аминогруппу на пировиноградную кислоту с образованием аланина, а сам превращается в гидроксипировиноградную кислоту. Последняя при участии НАДФН восстанавливается в глицериновую кислоту. Она переходит в хлоропласты, где включается в цикл Кальвина и образуется 3 ФГА.
|
||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 1146; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.23.92.50 (0.009 с.) |