Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Посттетаническая пластичность:Содержание книги
Поиск на нашем сайте потенция и депрессия.
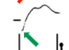
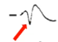
Процесс потенциации закл в том, что при некоторых воздействиях нейрон повышает свою чувствительность к естественному воздействию. Пример из работы Абрамовой. Она защищала кандидатскую у Пивоварова. Что тут изображено? Это помеченный известный нейрон. Это левый париетальный командный нейрон виноградной улитки, который называется «а3» (париетального ганглия). Слева изображён артефакт от электрич стимула. Стимул подают электрическим образом на афферент (а можно нейрон раздражать напрямую). После артефакта виден ВПСП, который генерирует нейрон. Этот ВПСП достаточно продолжительный (длится почти 2 сек). Вертикальные красные стрелки – это А. Если применим ритмическое раздражение, с относительно высокой частотой (см на фрагмент записи длительностью 2 мин), нейрон тоже будет генерировать ВПСП, кот сопровожд-ся иногда спайками. И по окончании стимуляции нейрон нах-ся в относительно депрессивном состоянии, поскольку высокая частота для нейрона не характерна. Через некоторое время можно еще раз нанести одиночное раздражение (рис С), как и в первом случае (рис А). Нейрон покажет повышенную активность. Как минимум в 1,5 раза А ВПСП увеличилась. Феномен потенциации сост в том, что после мощного воздействия нейрон повысил свою чувствительность и генерир-ет повышенную ВПСП.
Что же стало с токами? ВПСП – это потенциал. А нам важно, что возрастают токи, котор текут через мембрану. Методом фиксации потенциала можно этот нейрон зафиксировать и регистрировать токи. Эксперимент повторяется. Дается первый стимул, получаем ток (самая левая картинка) (около 20 наноампер (нА) у этого тока). Потом дают ритмическую стимуляцию (такую же, как и в предыдущ случае) в течение 2 мин. И после этого наносят точно такое же раздражение. Получаем увеличенный ток.
*Если ВПСП – потенциал, *Если ВПСТ –возбуждающ постсинаптич ток.
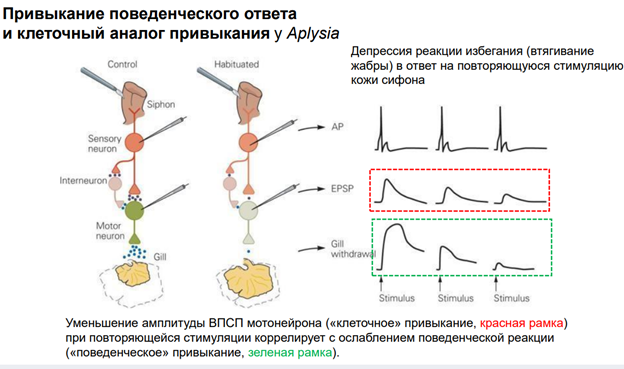
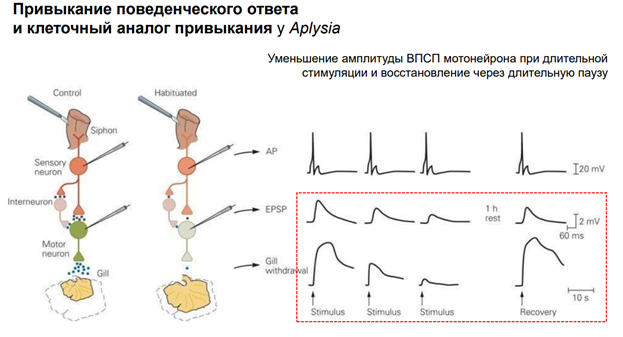
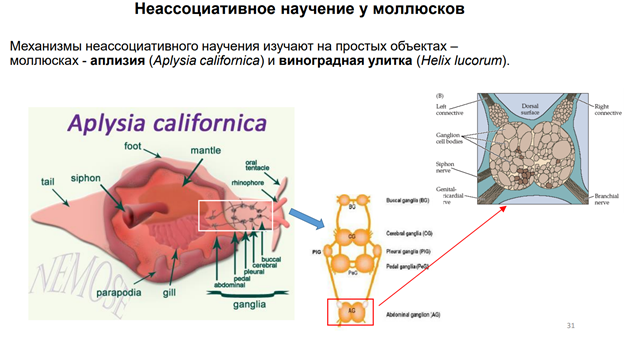
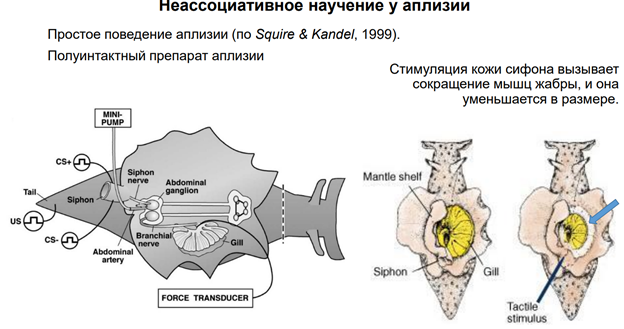
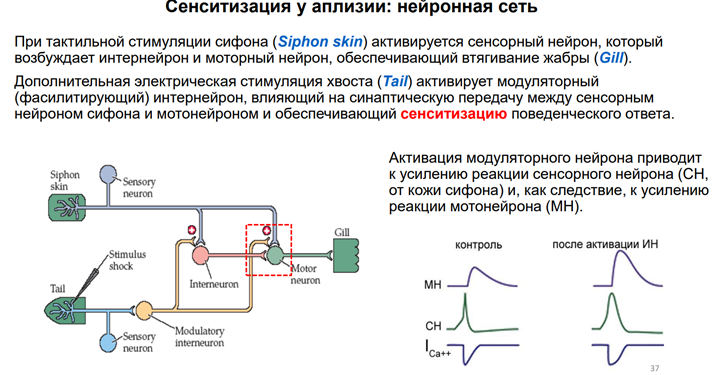
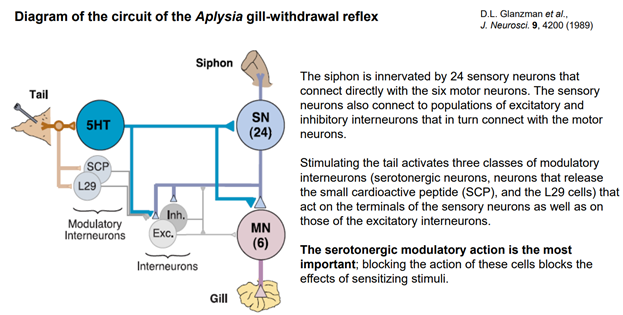
Эрик Кэндал и Гланзман. НС моллюска (аплизии) представлена 5 парами ганглиев. Эти ганглии содерж несколько тысяч нейронов. Это не так много по сравнению с НС позвоночных. Справа (голубой) нарисован ганглий (так рисуют в хэндбуках). В нем идентифицируются нейроны: сенсорные, моторные, … В нашем случае изображен абдоминальный ганглий. Он регулирует следующую прослой поведения – адаптивное. Быбрана такая поведенческая модель. Есть орган дыхания – сифон. Если его как-то потревожить (напри, уколоть или дать сюда электрический ток), то аплизия отвечает оч простым поведением – просто втягивает жабру. И это действие можно легко зарег-ть (его и визуально видно, но для колич оценки применялись приборы, рег-ие изменение). У виноградной улитки немного др поведение.
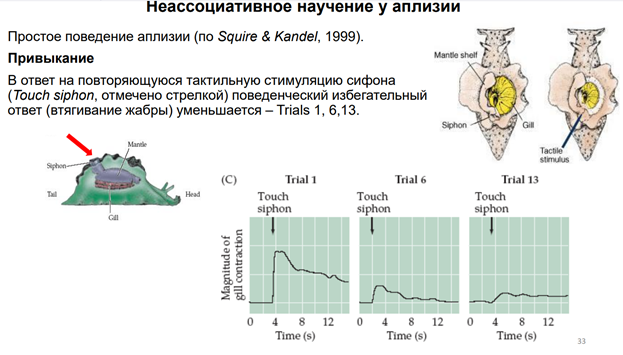
Тут показаны основные элементы оборудования. Force transducer - это прибор, котр оценивает силу (на сам деле срабатывает мышца (gill), и это усилие можно зарегистрировать). Тут есть некая ассоциация с классич павловским реф-сом. Хотя тут конечно не ассоциативное научение. Раздражение сифона считается «условным» стимулом, а раздражение хвоста – «безусловный». Для читстоты эксперимента исп-ся условынй стимул «+» и условн стимул «-». Это означает, что стимуляция сифона («+») сочеталась с электрическим раздражением хвоста. А другая стимуляция не сочеталаст, поэтому она названа «-». Т.о. «+» (там, где сочетали) был «потрепляемым», а стимул «-» - непродкрепляемый (индефферентный). У этого животного есть НС, кот обеспеч связь этих всех событий с активностью мотонейрона. А мотонейрон обесеч сокращение жабр. Феномены, которые получили: Если стимулируем сифон (тактильный стимул), получ уменьшение жабр, которое мож измерить. Что произойдет, если мы сделаем это много раз?
Первое прикосновение к сифону вызывает р-ю, изображенную на левой зеленой картинке (Trial 1). К 13ой попытке р-ия вытормаживается. Эта динамика демонстирует нам привыкание. Поведенческий ответ в виде втягивании жабры уменьшается.
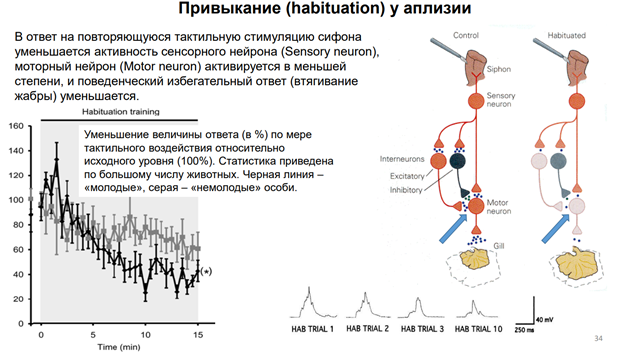
Если проводить это длит время и получить это на большом кол-ве животных, мож увидеть динамику (серый график). По оси ОУ – А ответа, по ОХ время в мин. Тут две кривые. Серная линия – нейроны моложых аплизий. Они быстрее адаптир-ся. А серые – пожилые – медленнее адаптир-ся. Чем объясн-ся феномен привыкания? Рассм-им нейронн сеть (картинка справа). Сифон касалочкой активируем. Сенсорн нейрон, кот обеспечивает восприятие этого дейстивя. Сенсор нейрон оканчивается напрямую на мотонейроне и на двух, по крайней мере, промежуточ нейронах (клетках, кот добавляют эффект сенсорного нейрона на моторн нейрон). Выдел-ся сначала много медиатора из сенсорного нейрона (глютамат). На моторном нейроне у аплизии на постсинап мембране есть к нему рец-ры. Много Глю, много рец-в активируется, мотонейрон дает мощный разряд импульсов. В нервномышечный синапс выдел много Глю. Мышца сокращ, уменьшает мышцу в размере. По мере привыкания из сенсор нерона выдел меньше нейромедиатора, след-но, на мышцу меньше, меньше сокращ мышцы. Это позвол-ет экономить ресурсы (привыкание).
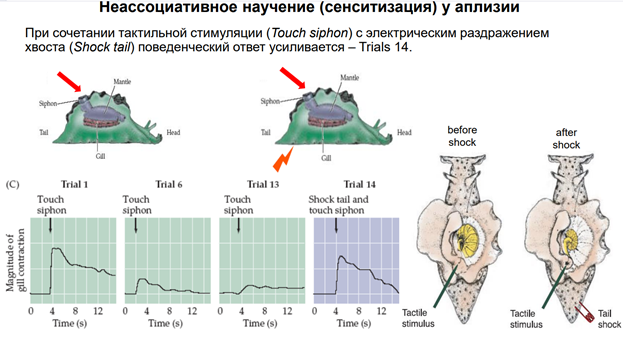
Что если привыкла нервная сеть. Если дадим раздражение хвоста (шок). Что произ поведенчески? См на синий квадрат. Жабра снова сократилась.
В зав от раздражения хвоста по силе зав результат на поведен уровне. Картинка слева. Тут по Ох часы: Контроль: Если не даем раздражение хвоста, то нейрон привыкает. Если даем один стимул хваоста, получим возрастание А в 1,5 раза, а затем снова угасание. Картинка справа. Тут по Ох дни. No shoсk – контроль – угасание р-ии Если 4 простых одиночных стимула – в 1 день увеличение, но к 4 дню адаптация Если 4 пачки – возрастание продожается долго, пости неделю Если по 4 пачки в день 4 дня, то эффект долгий и сильный (ответ в 8 раз усилится) Такая динамика напомин эффект долговременной памяти.
Это обобщ схема. Тут рег-ли активность отдельн нейронов. Сенсорного и моторного. Дендриты сенсор нейрона простираются в кожу сифона. При касании кожи сифона сенсор нейрон активир-ся. Этот сенсор нейрон дает аксон с колатералью на моторный нейрон. Т.е тут есть и прямая активация, и активация в обход (через переключение). Все события, кот обеспеч длительн эффекты, заключаются во фрагменте, выделенном на картинке красным пунктирным квадратом. Тут 2 окончания: 1) одно прих от хвоста, через кот мы осущ тетанизацию, 2) др прих через интернейрон от сенсорного нейрона, 3) но самый главн синапс – это между сенсор и мотор нейроном (голубой на картинке, тот, который напрямую идет). В эксперименте проводили внутриклеточную регистрацию. Контоль тут: р-я сенсор и мотор нерйона в отс тетанизации (в норме - без воздействия) Нам тут интересно, как ведет себя сенсор нейрон. Зеленым графиком обозначена его тетанизация, ВПСП (пик вверх или спайк). В ответ на возбуждение (на один спайк сенсор нейрона) мотонейрон отвечает ВПСП. Этот ВПСП обуславливает А р-ии. Т.е чем больше ВПСП, больше его продолжительность, тем больше выражена реакция. Тут еще кальциевый ток есть (нижний график). Это кальц ток в сенсор нейроне. Он не очень продолжительный с не оч большой А.
После активации. Сенсорного нейрона от хвоста модуляторный нейрон получ активацию, и простирается в область, выделенн красным пунктиром, образует аксо-аксональный синапс на пресинапсе от сенсорного нейрона от кожи сифона. Что видим на графиках? Сенсор нейрон демонстир более продолжит активацию – длинный ВПСП, кот мож зав-ся спйком. Моторный нейрон тоже усилив свой ВПСП тоже (т.к тут прямая синаптич связь). Кальциев ток, котор леж в основе такого события, более продолжительный. Это приводит к тому, что мышца жабры сокращ в большей степени.
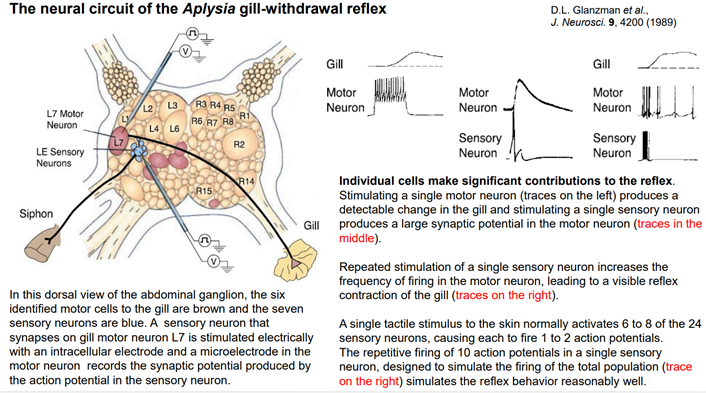
Оригинальн работа Глазмана. Он работал на Аплизии. Слева изображен ганглий. По дендриту от сифона происх активация сенсорного нейрона (тут он голубая фигулина) (на сам деле их тут несколько, поэтому и фигулин синих несколько). Моторный нейрон (красный L7 на картинке) располаг недальнего от сенсорного. Он простирает свой аксон к мышцам жабры (Gill). Тут еще обозначены рад нейронов, они известны. Каждое обозначение – L6, L3, R7, R1 – это все отдельные нейроны (не группы!). R означает, что это нейроны справа, L – слева. Это полуинтактный препарат. К интересующим нас нейронам подключены регистрирующие внутриклеточные электроды. (Если применим методику фиксации потенциала, то можем токи регистрировать). В данном случае через двуствольную пипетку (направлена к моторнейрону и сенсор нейрону) мож подавать стимуляцию (активир разл параметрами тока и получ ответ).
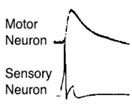
Если стимулир моторный нейрон, получи ответ жабры:
Если стимулируем сенсор нейрон, получаем активацию мотор ного нейрона:
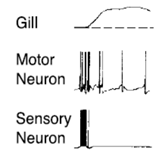
Если стимулируем сенсор нейрон более высок частотой, получм не просто ВПСП, а еще и спайки. Сначала частота спайков высокая, потом урежается. Это приводит к увеличеной р-ии жабры.
Это схема Сенсорных нейронов идентиф-но 24 шт. Они не всегда все активир-ся, это зав от активности стимуляции, площади поверхности активации (мы в сифоне одну точку активировали, а можно больше и сразу). Моторных нейронов, выходящих аксонами на жабру, 6 шт.
Тетанизируем хвост. Всего три нейрона реагируют на стимуляцию хвоста. 1) серотонин эргический нейрон (5НТ). Это модуляторный нейрон, кот вовлечен в сенситизацию оборонительн р-ии – втягиван жабры. 2) Серотонин эгрич нейрон (SCP), кот выдел из себя при активации малый кардиоактивный пептид. 3) L29 (малая клетка). 5НТ напрямую активирует сам сенсорный нейрон и посыл колатераль на пресинапс. SCP и L29 активируют некие промежуточные нейроны (Exc и Inh). Но самый главн модулирующий аксон – это колатераль от 5НТ к мотонейрону жабры.!
Тут некая феноменология. Перед сенситизацией мы на стимуляцию сифона получаем спайк в сенсорном нейроне (верхний левый спайк). Моторный нейро н дает ВПСП (EPSP), кот вызыв сокращение мышцы жабры и рефлекс избегания. После тетанизации через хвост (сенсор нейрно – фасилитирующий нерон – сенсорный нерон и колатераль на пресинапс сенсор нейрона (см картинку)) происх некие события (разберем позже), кот прподят к увеличению выброса медиатора из сенсор нейро на и больш активация мотонейона. Сам по себе сенсорн нейрон не сильно увеличив свою активность. Но при этом из него выдел больше медиатора (Глютамат-эргический синас), и мотонейрон демонстрир более сильн р-ю и по А, и по продолжительности. Это приводит к увеличен поведенч ответа. После тетанизации (после стимул-ии хвоста) (в отл от контроля) р-я нейрона сенсорного увеличилась, но не сильно.:
Привыкание приводит к тому, что уменьшается кол-во выделившегося медиатора. Мотонейрон и Сенсор нейрон выделяют Глютамат. Первые р-ии сильны, жабры уменьш в размере сильно, но в конечном счете, спустя 10 примерно стимуляций жабры сокращ меньше, хотя р-ия сенсор нейрона одинакова, поведенческий ответ прак исчезает. Если сделать паузу примерно в 1 час, не трогать, то р-я примерно возвращается.
В этом эксперименте (это все мы про аплизию, ее сифон и хвост) был проведен контроль. Проверяли, связан ли во времени активация сифона и стимул-я хвоста. Если эти 2 события разнесены во времени в случайном порядке (как в услов реф-се сочетание УС с БС – тут это, конечно, в кавычках, слишком уж простой организм аплизия) (см картинку под номером 1), то не увидим никакого эффекта: увеличения ВПСП на мотонейроне. В классической еще школе на поведенческом уровне было показано, что Если БС и УС не сочетаются во времени, то УР нет. А тут то же показали на клеточном уровне! Каеф! Вывод: при непарной стимуляции сеситизация ВПСП мотонейрона не происходит.
А что будет при парной стимуляции? Даём раздражение сифона («условн» стимул) и подкрепляем его раздражением хвоста («безусловный» стимул). Интервал 5 мин Парная стимуляция дает сенситизацию ответа мотонейрона и, соотв-но, поведенческого ответа.
Оригинальн работа Кэндола. Показана реальн запись в эксперименте. Фрагмент А (верхний правый угол) 1: тестовый стимул – малая А ВПСП мотонейрона, 2 и 3 – тест, а затем раздражение хвоста (сенситизация = прайминг тут = тетанизация). После тетануса дают обычный стимул на сифон – получаем увелич р-ю,… потом опять тетанизируем. Потом пауза в 10 сек – 4 картинка. Потмо смотрим что происх через 3 мин, 10 мин, 20 мин…
Если отложить эту феноменологию на график (Ох- минуты, Оу- проценты). На 6 мин тетанизация произошла. Буквально 2 мин уходит на возрастание р-ии в процентах от контроля (тут А ВПСП), а потом по экспоненте уменьшение, и в конце концов выйдет к изначальному уровню. Нижний самый график (плоский почти) – мембранный потенциал, котор сопровождает облегчение А-ы ВПСП. Т.е. в мотонейроне этой длительной стимуляции немножко увеличивается потенциал (ПП). Нейрон стан-ся деполяризован примерно на 6 миливольт. Почему? Объяснение в этой работе не дается. + в 1965 году не знали о метаботропных рец-х, ионные тоже были прохо изучены. Если вспомнить Общую нейрофизиологию: из чего скалад общая активность нейронов? Кроме потенциалзависимой проводимости ионотроп и метаботропных каналов есть еще транспортные с-мы, котор усиленно работают, и их оч много.
Оригинальные записи. По сравнен с предыд рисунком палитра инвертирована (запись с осцилографа). Предыд слайд из хэндбука.
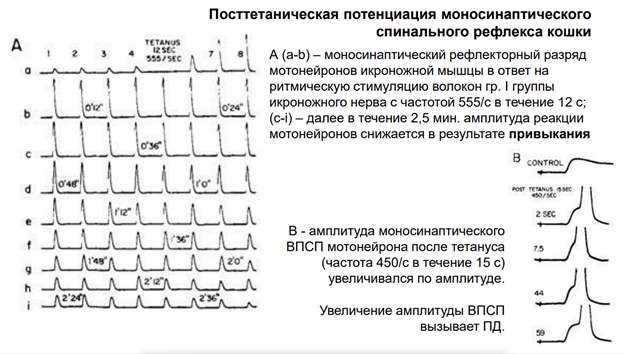
А что же на позвах? Примерно в то же время Джон Эклз показал многую феменологию на нейронах СМ (спинного мозга). Одни из первыз мотонейронов были записаны на СМ. Тут приведён пример посттитанической потенциации моносинаптического спинального реф-са кошки. Тут приведены записи на а и б – моносинап рефлект разряд группы 1 – афференты идут из мышцы, интрафузальные волокна - рецепторы растяжения. Это методика Эклза. Эти афф-ты были идентифицированы, их можно было стимулировать электрич способом с высокой частотой (555 импульсов/сек). Потенциировали в теч 12 сек. При этом получили увеличение А ВПСП (шкала временна немного сжата). Этот эффект длится довольно долго: На фрагменте с с до i он все еще есть, хотя А снижается. В итоге, примерно через 2,5 мин доходим до исх уровня.
Такую потенциацию мож увидеть на др нейронах в др местах. Св-во потенциироваться и привыкать им прак все нервные элементы.
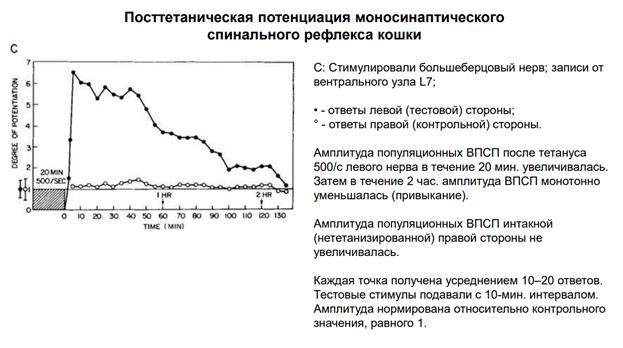
Тут 20 мин - референсные значения получались. Обеспечивали потенциацию тетанизацию с одной стороны СМ и с др стороны. Т.е тут 2 мотонейрона – тот, котор потенциировался, а на др стороне тот, кот (для контроля) стимулировали одиночным стимулом, чтобы посм, будет ли меняться его активность. Длительность эксперимента более 2 ч. После тетануса левого нерва в течение 20 мин 500 имп/сек (черный график) одиночный стимул давал увеличение А. И в теч 2 ч это все уходило к нормальному сост. А вот А не тетанизированной (правой стороны) не приводило к увелчиению А. Кажд точка – усреднение ок 10 ответов. За единицу (ось Оу) были приняты референсные значения.
До сих пор рассмотрели мех-м пластичности, кот происх на пресинапсе. Тетанизация происх благодаря соб, происх-им на пресинаптической терминали.
Сейчас постсинаптическую пластичность рассмотрим и долговременную потенциацию в гиппокампе.
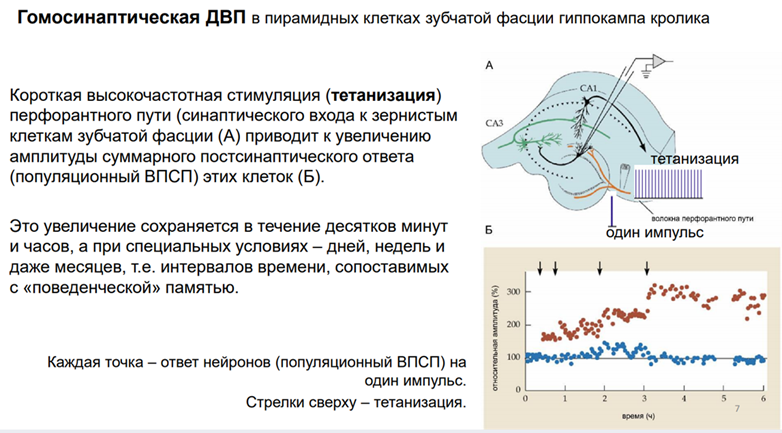
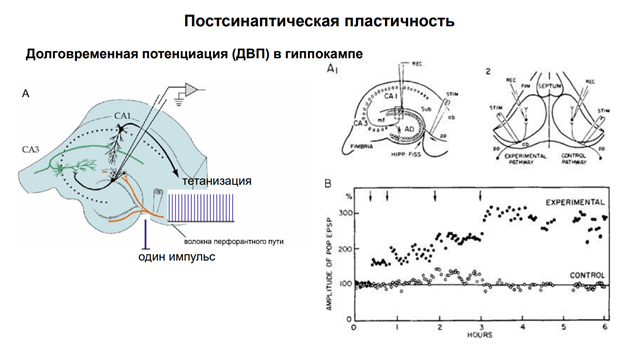
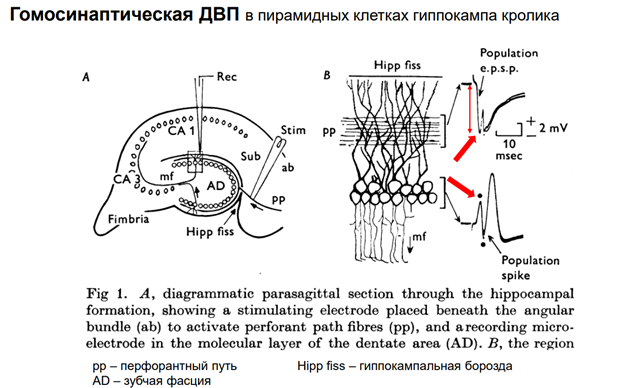
Первый объект, на котором получена долговременная потенциация – гиппокамп. Гип-п отвеч за опред виды памяти. Срез гип-па. Если рег-ть нейроны зубчатой фасции, стимулир-ть перфорантн путь. После тетанизации нейроны зубчатой фасции на одиноч стимуляцию того же самого пути демонстрир повышенн ВПСП.
Гомосинаптическая ДВП
Тери Лёмо. Первооткрыватель феномена долговрем мотенциации. Его работа - первая работа, посвящ долговременной потенциации.
Долговремен потенциация им отношение к пластич перестройкам, его мож рассм как клеточ аналог долговременной поамяти. Тут показана схема эксперимента. Регистрировали нейроны Зубчат фасции, стимулировали перфорант путь. Феномен был получен не на срезах, а на мозге. На кролике. У него большой гип-мп (по сравнен с мышами и крысами). Регистрировали суммар активность гип-ого слоя. Надо регистрировать именно активность пирамид!! Что и было сделано в этом эксперименте.
Тут изображен слой пирамид, вверх простираются дендриты:
PP – это волокна перфорантного пути. Это афференты – входы нейронов Зубчат фасции. Что регистрировали по слоям?? Если электрод поместить в область дендритов, то имеем популяционный спайк – внеклеточ активность на дендритов нейронов. Отклонение вниз будет.
Если погрузим электрод в слой пирамид, получим популяционный спайк. Отклонение наверх будет.
Обычно при этом регистрир-ют А этого спайка от одной точки, до другой (точки видны на последней картинке) – это называют горка. Она отражает первичный ВПСП пирамид в ответ на стимуляцию волокон перфорантного пути. Остальное – это уже не ВПСП – другие, вторичные события А несколько миливольт – большая. В эксперименте измеряют или А, или фронт.
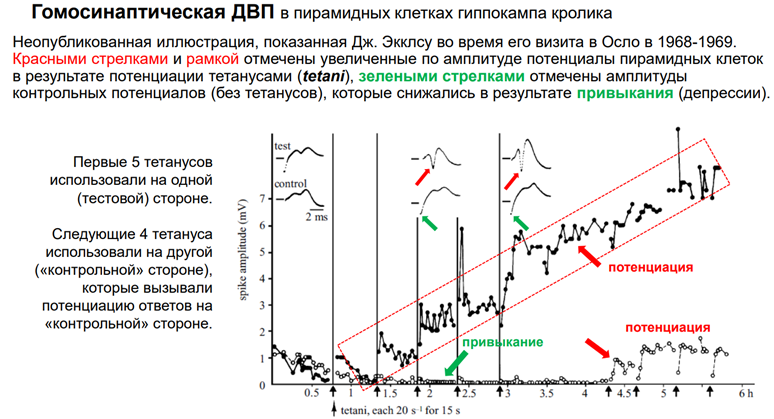
Мы привыкли, что с той статьи Лёмо все и началось. Но нет. До этого были еще три статьи (его же 😊). А началось все с одной картинки (см выше). Для научника Лёмо все это не оч было интересно, поэтому грустно, ибо Лёмо был аспирантом. И пока туда не добрался Близ (Бриз?). А что это за рисунок? Это результат эксперимента. Его Лёмо показал Джону Эклзу его. И Эклз сказал, что это открытие, надо публиковать. Эксперимент. Черные точки и прозрачные – это 2 гип-па. Один с одной сторны, др – с др. Потенциация осущ только с одной стороны. Это одиночные стимулы (электрич стимуляция) через перфорантн путь. Регистрир-ся популяционный спайк в нейронах зубчатой фасции. Наблюдается феномен привыкания. Т.е если давать неск стимулов подряд, то ВПСП уменьшается по А. Причем уменьшается как на своей стороне, где мы делаем тетанизацию, так и на противоположной, где тетанизации нет. Вертикальные полосы черные – момент тетануса. С частотой 20 стимулов /сек в течение 15 сек даетм тетанус. Т.е. дается пачка 300 стимуляций за 15 сек. Каждый последующий тетанус даем через пол часа. И вот до первой вертикальной полосы (до тетануса первого) гип-п уже привык – там динамика на спад идёт. А дальше – на той стороне, где нет стимуляции, продолжается депрессия ответа после одиночного стимула. А где была тетанизация – тенденция к возрастанию. Но при этом внутри каждого промежутка есть привыкание. Но каждая последующая тетанизация приводит к увеличению А. После 5ой тетанизации наблюдаем длительную восходящ-ую динамику – потенциация. Причем потенциировался наш ВПСП в 8 раз, если сравнивать с контрольн уровнем. На контрольной же стороне не наблюд увелмичения А. Вконце дале 4 тетануса – и получили тоже тетанизацию, просто не таку. Выраженную. Если посмотреть на единичные ответы, то в контрле они маленькие, без тетанизации:
А в тетанизированной стороне происх увеличение А (поп спайк увеличивается по А):
Т.о. была показана сп-ть нейронов Зуб фасции увеличивать свою активность.
Несколько слов про технологию. Перешли уже на среды – меньше надо животных. Много получить много средов – сотни даже. Эти срезы можно положить на подложку, котор им сетчатую основу. И срез мож долго сущ-ть (дни или недели), если доб растворы, агенты, перфузировать… И вот на этой сетчатой подложке изображен гип-п, видны его листки. Темные полосы на нем – скопления тел пирамид. Под бинокуляр микроскопом можно подводить в эти слои электроды.
На этой схеме потор эксперимента Тири Рёмо. Но на одиночных срезах. Регистрировали популяцион спайки нейронов Зубчатой фасции. Стимулировали перфорантн путь одиночным стимулом. Если дать тетанизацию однократную с высокой частотой в течение нескольких сек (протокол мож быть разный на сам деле). Что происх с поп-спайком (популяционным)? (каждая точка – это одиноч спайк популяционный, ось Оу - за 100% взяли в началае – в норме, Ох – время в часах) В контроле (если ничего не длать): ничего не происх. А если тетанизировать через опред промеж вермени (время в часах), после кажд тетанизации получ увеличение А ступеньками. После 4ой тетанизации уже 300% - А спайка увеличилась в 3 раза. Долго держится причем. Такие долговременн эффекты ассоц-ся у нас с долговременной памятью. Это протокол гомосинаптической долговременной потенциации. Гомосинаптическая – это значит, что мы и потенциирует данный вход (эффекрент), и через него же проводим одиноч стимуляцию.
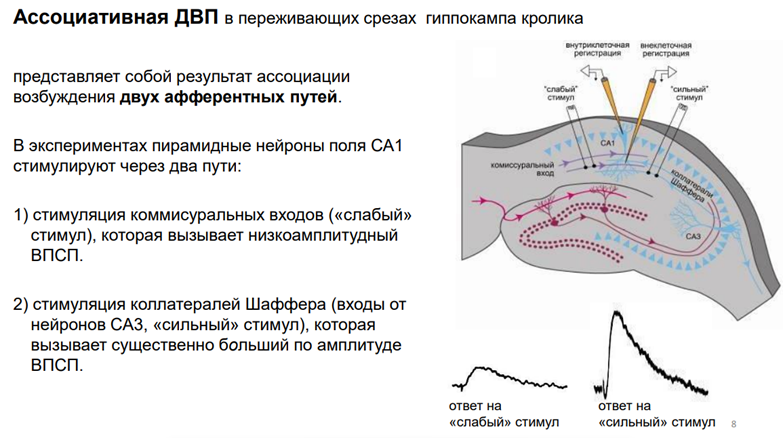
Хочется, конечно, проинтерпритировать, как в сл сенситизации на моллюсках в павловской терминологии воспроизвести протокол с услов и безуслов стимулом (УС и БС). Это тоже бвло сделано на гип-пе. Тут уже парная стимуляция. Эта долговременная потенциация уже наз-ся гетеросинаптическая (ассоциативная). А раз ассоциативная, то возникает представление об условном реф-се. Эта ассоциативная ДВП представл собой результат ассоциации возбуждения двух афферентных путей. В этом эксперименте нейроны поля СА3 (синие пирамиды), их дендриты простираются в глубину. На этих дендритах оканчиваются синапсы от 2х входов: 1) связи комиссуральных волокон (комиссуральн вход) – связывают поля гип-па; стимуляция вызывает низкоА ВПСП, 2) стимул-я колатералей Шаффера, кот идут из Зубч фасции– оканч синапсами на дендритах пирамид – сильный вход, т.к при его стимулиции возникает высокоА спайк (поп-спайк). Элктроды слабого и сильноко пути – мож регистрировать события на дендритном дереве, а можно осущ внутрикл регистрацию пирамид (получать активность одиноч нейронов).
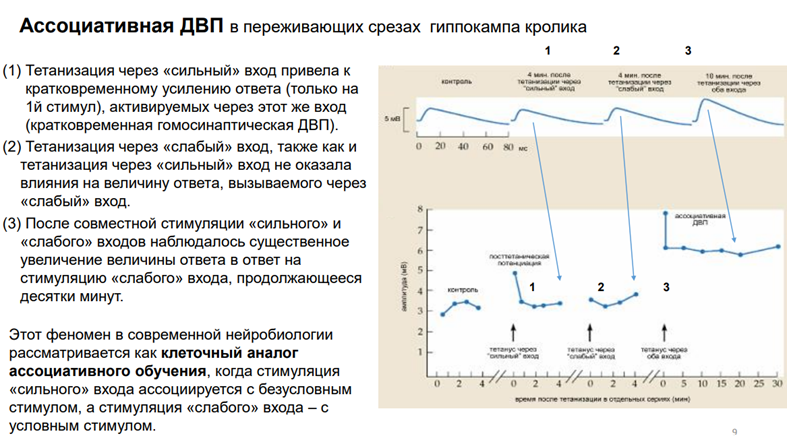
В хэндбуках так отвечают. На этом графике (верхний) три фрагмента. Это одиночные записи, это ВПСП, которые рег-сяв нейронах поля СА1. А на нижнем графике изображены некие события, феноменология А-уд поп-спайка. В контроле ничего не происх. Наносим одиноч раздражение через слабый вход и получаем 3 миливольтовые поп-спайки. Тетанизация через «сильный» вход – кратковременное усиление ответа только на первый стимул, активируемый через этот же вход. Т.е мы тетунасом простимулировали «сильный» вход и через него же дали потом одиноч стимул. Получили ответ увеличенный, но только на первый стимул, а все остальное к норме вернулось. Тетанус на «слабый» вход. Не получили на одиночный стимул никакого увеличения. Тетанус через оба входа. Существенное увеличение А в первой стимуляции. Но в данном случае повышенная А осталась как минимум в течение получаса.
Модель классического УР. Продемонстрирована на примере ассоц-ой долговрем потенциации на нейронах гип-па. *«слабый» - с уловным ассоциируется, а «сильн» - с безусловным стимулом. А – тетанич стимул-я только через «слаб» вх не вызыв долговрем изменений Б – тетанич стимул-я через «сильн» вх вызывает увеличение эффективности «сильного» синапса (это пример гомосинаптической ДВП). Получили потенциацию гомосинаптическу., но при этом не получили ничего, когда стимулир слабый вход. В – оба входа стимулируем. Тетанизация, после него и там, и там ответ (гетеросинаптический ДВП). Эти события ложатся в схему классического Павловского реф-са. Но мы говорим, что это клеточ аналог.
Посттетаническая депрессия
Кроме потенциации есть еще феномен постсинаптической депрессии. Это было сделано Эвартсом. Потенциация – это процесс в одну сторону, а депрессия – в др. Тут показан постсинаптический ответ латерального коленчатого тела после тетанизации контрлатерального оптического нерва у кошки в течение 20 сек. Исходные ответы были приняты за единицу. В течение длительного времени (почти 3 ч) наблюд депрессия потенциалов.
Феномен долговрем потенциации получаетчя прекрасно на разл участках коры ГМ (головного мозга), подкорков структурах. Это свид-во о том, что это универсаль мех-м, кот свойственен для многих синапсов.
Привыкание
Привыкание поведенческих ответов. Клеточн аналог привыкания
Костистые рыбы, включая Данио рерио, демонстрируют простую двигательную реакцию на испуг, которая контролируется двусторонней парой больших командных нейронов, клеток Маутнера, в заднем мозге рыбы. Двигательная реакция (C-start) запускается резким сенсорным (слуховым, визуальным или тактильным) стимулом. Эта реакция была зарегистрирована у личинок Данио на слуховой стимул через 4 дня после оплодотворения. Причём также было выявлено привыкание к стимулу. Функция данного поведение – избегание хищников.
Были описаны три формы привыкания: 1) Быстрое привыкание. (рис Б) Частые повторы стимулов – 50-120 шт за 1 мс (200Гц). После завершения тренировки через паузу в 3-15 мин поведенческий ответ возвращается к тому, котор был до привыкания. 2) Кратковременное. (рис А) Эффект сохраняется до 1 ч после тренировки. Предъявление стимулов блоками (всего их 10) по 900 шт (1Гц), интервал между блоками 5 мин. Было обнаружено, что для данного привыкания необходима активация N-метил-d-аспартата (NMDAR). 3) Длительное привыкание. Стимулировали 6 блоками, разнесённые во времени; в каждом блоке по 8 коротких слуховых стимулов (1 Гц). Привыкание сохранялось до 18 ч. Индукция данного привыкания также зав-т от NMDAR.
Что по рисунку Б: возврат к предтренировочному уровню через 15 мин. Звездочка указывает, что 10-секундный тест значительно отличался от предварительного теста и 15-минутного теста. Решетка – указывает, что 1-минутный тест от предварительного теста и 15-минутного теста.
* Сенсабилизация – форма неассоциатривного обучения, при которой воздействиее возбуждающего стимула вызыв усиление реакции. Если мы этим же стимулом усиливаем привычную реакцию – это дезабитуация ( DISHABITUATION).
Интересно, что введение DL-2-амино-5-фосфонопентановой кислоты вместе с никотином блокировало сенсибилизацию. Следовательно, активность NMDAR, видимо, опосредует некоторые формы привыкания и чувствительности.
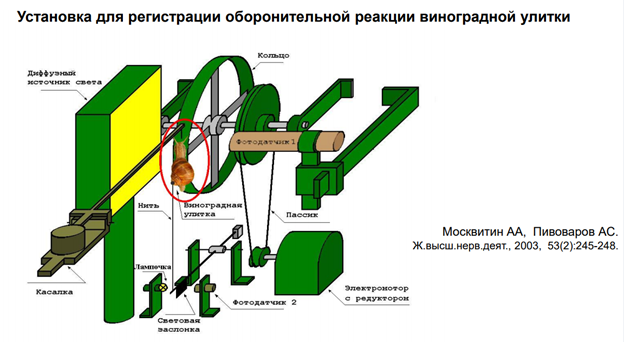
Объект: морская улитка Aplysia californica. Привыкание - это снижение реакции на повторное применение данного стимула, которое нельзя отнести к сенсорной адаптации, сенсорной или моторной усталости (Thompson and Spencer, 1966; Rankin et al., 2009). Интенсивность стимулов (последовательности 500 мсек [25 Гц] импульсов тока 10 мсек) устанавливалась для каждого препарата выше порогового значения для надежного вызова отдергивания жабр и оставалась неизменной на протяжении всего эксперимента. Схема эксперимента: Стимулировали сифон – две стороны: контрольная и стимулируемая (=тренированная). Предтренировка: Стимул в тренируемую сторону, через 5 мин в другую (контрольную) сторону. Ждали 1 час, потом эксперимент. Всего было 5 блоков стимуляции. Интервал межблочный = 90 мин. Во время каждого блока тренируемой стороне давали 30 стимулов. Далее после 2 ч отдыха обоим сторонам давали по 1 стимулу (с разницей во времени 5 мин). Результаты: Тренировка привела к долгосрочному привыканию рефлекса отвода жабр к стимуляции тренированной стороны сифона. Закрашенные кружки представляют реакцию на стимуляцию тренированной стороны, тогда как закрашенные квадраты представляют реакцию на стимуляцию нетренированной стороны.
Механосенсорные нейроны запускают высокочастотные импульсы в ответ даже на умеренные тактильные стимулы на кожу. Сенсорные нейроны также запускают вспышки в диапазоне 1-60 Гц в ответ на электрич стимулы. Метод. Электроды были 1 в сенсорном нейроне, 2 других (регистрирующий и вводящий ток) мотонейронах. Двигательный нейрон был гиперполяризован в теч всего эксперимента до -80мВ, чтобы предотвратить суммацию ВПСП от запуска ПД. Сенсорный нейроне стимулировали 1 сек при частоте 10 Гц (10 мс импульсы, 100 мс межимпульсн интервал) током, котор в 1,3 раза превышал пороговый. Далее между блоками использовался интервал в 10 мин, чтобы синапс мог восстановиться. Всего было проведено 10 стимуляций. Результат: Сенсорные нейроны запускают всплески ПД при активации механическими стимулами, приложенными к коже. Рис 1А: ответ сенсорного нейрона сифона на серию контролируемых механич стимулов на коже. Шаг между воздействиями 800 мс, кле точный разряд увеличивается в зависимости от интенсивности стимула. Рис. Б: хвост стимулируют, полуинтактный препарат. 200 сек стимуляция длится (ее начало обозначено стрелкой - shock). Сенсорный нейрон срабатывает с частотой, соответствующей стимулу (60 Гц), с небольшой адаптацией. За этим разрядом следует другой разряд, во время которого частота сот-ет 10 Гц.
На рис А: пример всплеска ПД, вызванного в плевральном сенсорном нейроне (SN) и результирующего суммиранного ВПСП в двигательном нейроне (MN). Амплитуды отдельных ВПСП снижаются до установившегося уровня 20% от начального ВПСП. Постсинаптический ответ был разделен на три фазы: инициация ВПСП, переходная фаза (со 2 по 5 зубец) (в теч которого синаптич эффективность снижалась) и фаза устойчивого состояния (с 6 по 10 ВПСП) (в теч которого амплитуды ВПСП достигли стабильного уровня депресси). Рис Б: совокупные данные для всплесков 10 Гц, в котор амплитуды ВПСП были нормализованы к амплитуде начального ВПСП.
В эксперименте проанализировали 40 клеток (14 ЛПа3 и 26 ППа3). Мембранный потенциал кл-ок варьировался от -45 до -85 мВ.
NCR – никотиновые рецепторы
MCR – мускариновые рецепторы
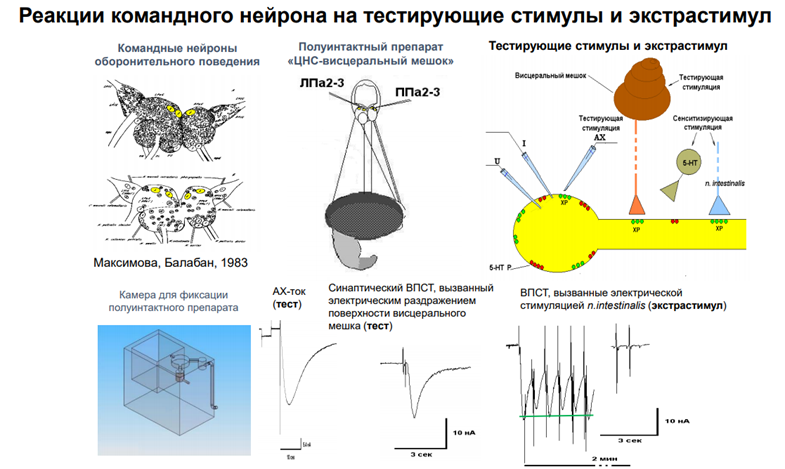
Анатомическая специфика. В экспериментах были идентифицированы эти синапсы. Они оказались холинэргическими. Были идентиф-ны холинэргические нейроны. И, соотв-но, рецепторы мембран этих нейронов. И для имитации таких возд-ий был использован искусственный синапс. К тому месту, где на мембране нейрона раса-сь АЦХые рецепторы подводилась пипетка, наполненная АЦХ. И методом микроионофореза мож было выделять из пипетки дозированное кол-во АЦХ на пов-ть нейронов. 1) Можно было менять концентрацией АЦХ в пипетке. 2) Мож было варьировать длительностью и интенсивностью тока для электрофореза. Чем больше ток, тем больше Ацх выделится. Т.о. иск синапс заменял реальный. На нейрон иммитировали тактильное воздействие кол-вом ацх, которым воздействовали. При этом были зарег-ны токи (методом фиксации потенциала). Надо было исп пипетки тонкие. Метод пэч-клямп целой клетки был, можно было фиксир потенциал на нуж уровне и рег-ть токи
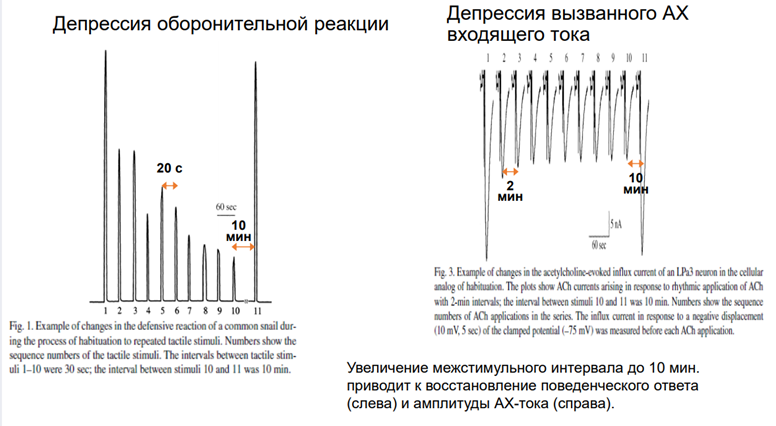
Картинка справа. Регистрировали токи. Интервал между воздействиями ацх-ом 2 мин. Токи входящего направления, т.к ацх рецептор обеспеч ВПСП, токи катионовые д.б входящими. На первую аппликацию ацх-ом получаем высокоА ток. Если через 2 мин повторяем стимул-ю, ток уже меньше. На 4 и 5 наступает плато. Если сделаем перерыв, получим восстановление А. Заключение: таким поведением управляет командные нейроны, актив-ся ацх-ом.
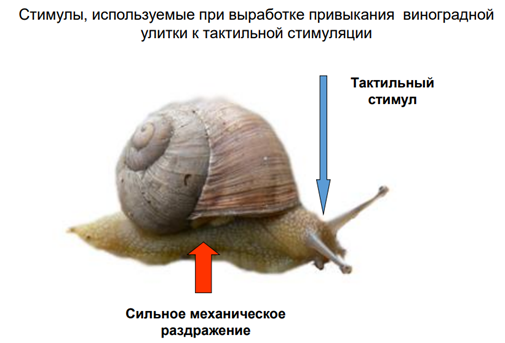
Чечь идет о сенситизации оборонитльной р-ии. При касании улитки за антенулы, она сожмется, но не до конца. А если проведём тетанизацию по хвосту, и после неё осущ-м тактильную стимул-ю, увидим р-ю большей А.
Тут есть картинка из работы Балабана. Командные
|
|||||
|
|
Последнее изменение этой страницы: 2022-01-22; просмотров: 118; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.2 (0.013 с.) |















 хотя вначале эксперимента он был такой же, как и контрольный.
хотя вначале эксперимента он был такой же, как и контрольный.