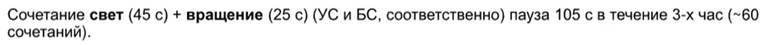Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Определение пластичности (полное)Стр 1 из 20Следующая ⇒
Пластичность нейронов. Феноменология Пластичность в целом – Майоров, пластичность нейронов – Пивоваров
Котляр основал на каф-ре нейрофизиолог мех-мы обучения. Его исследования были на высоком уровне (на мировом).
Опыт организм приобретает в пренатальном и потом в постнатальном периоде жизни. И далее в теч жизни. Нейропластичность заключается в том, что мозг может восстанавливать утраченные связи после различных повреждений и патологий. И мозг может изменять свой ответ на внешние воздействия (приобретённые формы поведения, виды памяти..)
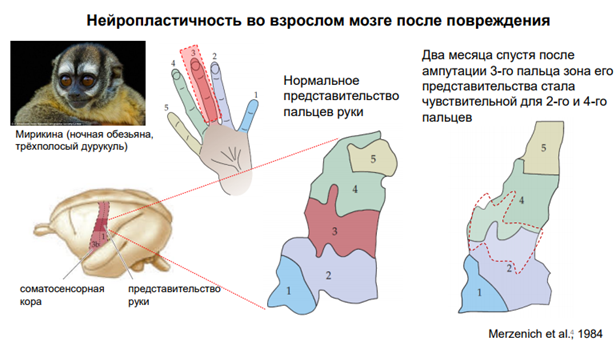
Это пример проявления нейропластичности во взрослом мозге после повреждения. Эксперимент проведён на обезьяне. В сенсомоторную кору проецируется в том числе и кисть, и занимает там большое место, т.к руки у обезьян играют важную роль в их поведении (Ну и в эволюции так было решено, что чем сложнее функция, тем больше нейронов нужно для обслуживания этой функции). На этом фрагменте соматосенсорной коры показаны области, котор соответствуют пальцам руки от 1 до 5. Тут зарегистрированы нейроны, которые отвечают за иннервацию кожных афферентнов, кожных рецепторов, которые сообщ-ют о некоторых воздействиях на эту кисть. Ампутировали третий палец. Прошло 2 мес, зона его представительства «растворилась», но нейроны не исчезли, они приобрели чувствительность к другим (соседним) пальцам – 2 и 4. На более правой соматосенсорной коре показан контур красный, где примерно была раньше зона для 3-его пальца. Т.е. нейроны не погибли. Они, конечно, потеряли прямую афферентацию. Но они перестроили свои синапсы и обрели чувствительность к сосед пальцам.
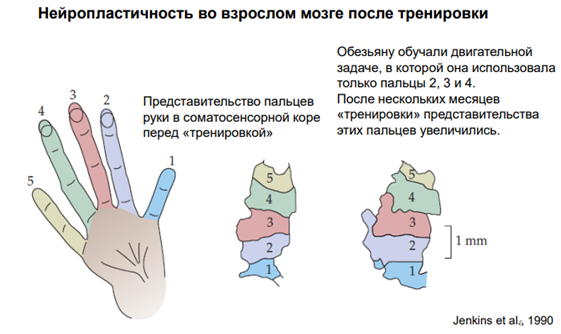
Другой – более щадящий эксперимент. Опять перед нами представительства пальцев в соматосенсор коре перед тренировкой и после. Обезьяну обучили некой двигательной задаче. 2, 3 и 4 пальцем надо было выполнять эту задачу (так задали экспериментаторы). После нескольких мес тренировки площадь поверхности соматосенсор коры для этих пальцев увеличилась. Прошлые связи нейронов остались, анатомически они никуда, по идее, не должны были деться. Можно предположить, что у старых нейронов атрофировались синапсы ненужные и возникли новые (синаптогенез). Т.о нейрон остается живым, но приобретает несколько иные свойства. И аксоны, и дендриты мог прорасти. Аксоны мог направиться к дендритам, сформировать синапсы. На молекуляр уровне: активируются факторы роста, которые приводят у синаптогенезу (возникновению новых синаптических окончаний).
Сформировавшаяся НС (нервная система) не стабильна, в результате взросления число синапсов уелич-ся, со старением уменьшается.
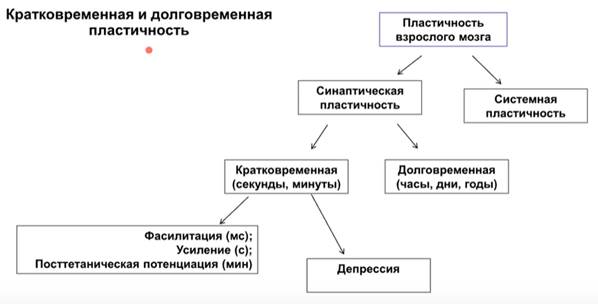
Пластичность нейрона. В резул перестроек нейрон может что-то усилить (потенциация) или ослабить (депрессия).
У нервных кл-к есть ещё такие функциональные характеристики, как перекачивание ионов против концентрационного градиента, что составляет возможность существования этих клеток. Нерв клетки пропускают по своим каналам ионы в обе стороны – основная и наиб специф-ая хар-ка. Конорски и Костюк. Они определяют пластичность как длительн модификации клеток и синаптич функции. Они говорят, что в основе модификаций поведения лежат перестройки в нейронных сетях. Основные феномены, кот отраж-ют изменение эффективности синапт передачи: · Длительн потенциация · Гетеросинаптич облегчение · Неассоциатив формы научения. В общей классификации памяти их называют стимулзависимое поведение. Оно не ассоциативное, т.к нам достаточно всего одного стимула, чтобы получ на него привыкание/ сенситизацию. Для сенситизации нуж опред условия (далее) · Ассоциатив научение, котор представлено рефлексами различной сложности – классический и инструментальный.
Эрик Кэндэл. Нобль лауреат. Обобщил многолетний опыт своих предшественников. С 60х работал в области синапт пластичности. Аплизия – большой моллюск. На нем изучали не ассоциативные формы научения. Написал книгу «Клеточные основы поведения», тут значит часть посвящ пластичности. Дэвид Гланзмэн. Работал
Наши соотечественники, котор внесли опред вклад в исследование пластичности. Соколов. Латанов работал с ним более 10 лет в области цветового зрения. Заложил основы в представлении... Он не занимался пластичностью напрямую, но он дополнил современными даннымимодель рефлекторной дуги (дополнил Павлова). Он был заведующим кафедры Психофизиологии МГУ.
Сахаров. Занимался беспами. Написал книгу «Генеология нейрона», где он описывает эволюцию развития мех-мов, кот опред пластические перестройки беспов. Есть значимые мания, как нейрон интегрирующ информацию, кот в него поступает. Работал на простом поведении моллюска (это хорошо, т.к. простые Нервные с-мы (НС) дают более однозначный резул-т, хотя более редукционистский). Он считал, что все дело в медиаторах. И пластичность опред-ся различными воздействиями со стороны разл медиоторных с-м. И вступал в полемику с теми, кто рассм ПД, как отражающие вопросы пластичности. Но и без Сахорова понятно, что ПД (потенциалы действия) – это способ распространения инфо на (относительно) далёкие расстояния. И сам ПД – все или ничего. И если «все или ничего», то у него два состояния – «ноль» и «единица». И как мож быть закодирована инфо, кот поступает в ПД? Никак! Может, если говорить о числе спайк ов, их продолжительности, паче и т.д. Но все, на сам деле, просих на мембранах нейронных кл-к, на синапсах. И синаптич паттерн активации и явл тем феноменом, кот отраж содержательные некие имения в мембране нейронов. Балабан. Пивоваров. Работал на отдельных клетках
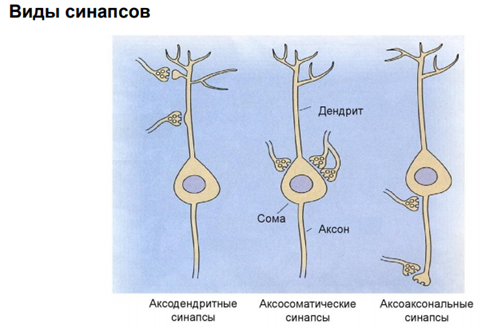
На сам деле такие сп-бы контактов – это такой сп-б разделить потоки инфо-ции, которая так или иначе влияет на состояние нейрона, его функционирование в сетях нейронов. Синапсы быв разные по их локализации, эргичности, морфологии…
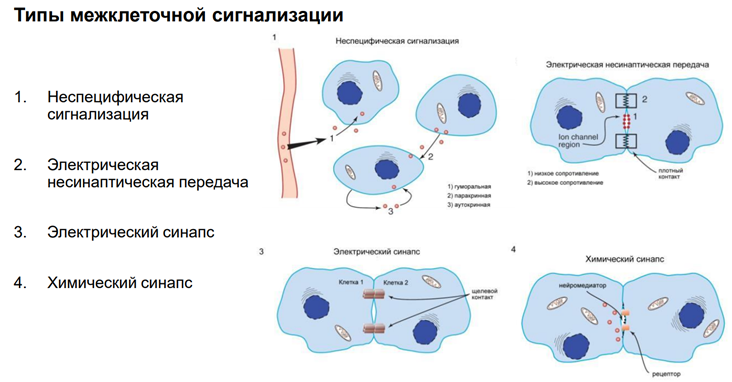
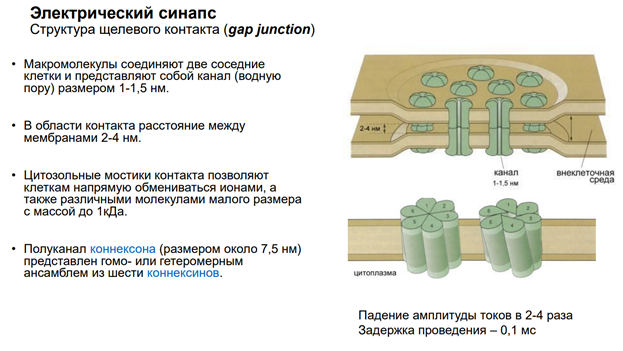
Мы в основном будем говорить про химический синапс. Но есть ещё неспецифич сигнализация. Когда агент, который влияет на нейрон, им происхождение не из аксонов, а из крови, неклеточной жидкости. Это гуморальная (паракринная и аутокринная) регуляция. Так действуют гормоны. Здесь неспецифическая связь. Эффект определяется только тем, есть ли рецептор на клетке к этому вещ-ву или нет. Электрич синапсы. Нам в этом курсе не интересны, т.к. синаптич перестроек тут не следует ожидать из-за того, что тут простые образования – представляют из себя «провода», котор соед-ют клетки. А химический синапс нам интересен, т.к. многообразие медиаторов / веществ, которые имеют нейромедиаторную природу, обеспеч разнообразные эффекты
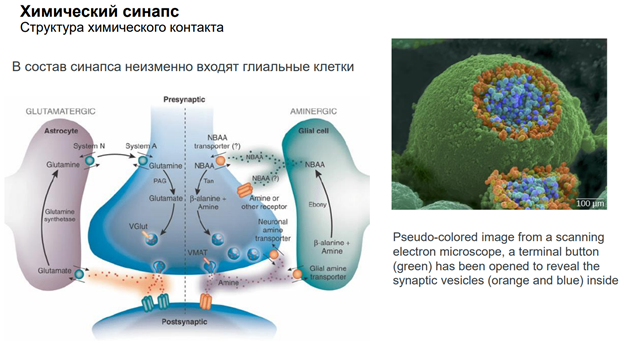
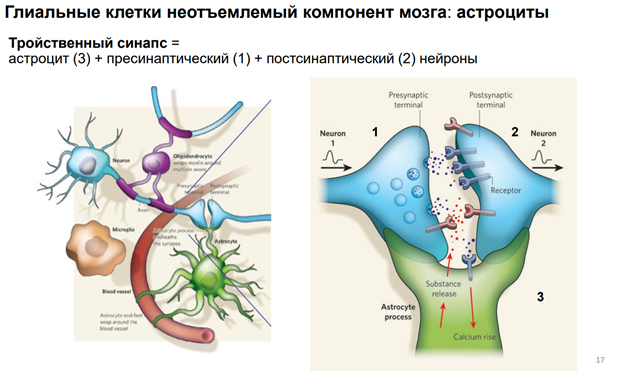
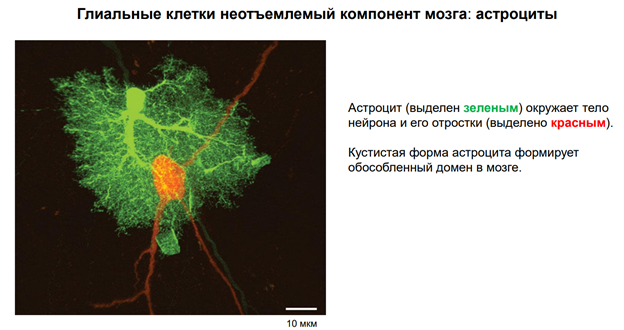
Глия. Сейчас к ней много внимания. В частности, к астроцитам, кот являются участниками синаптич передачи. На картинке показано, как астроциты окружают клетку. Астроциты бывают глутаматэргические и аминэргические. Они происх из общего предка в эмбриогенезе (из стволовой кл). Они ближайшие родственники нейрона. Астроциты имеют фактически все св-ва нейронов. Там есть и транспорт с-ма. Например, без астроцита не может осущ-ся глютаматэргическая передача, поскольку предшественники глутамата синтезируются в специальном астроците (розовый слева на картинке), и без него не могло бы быть никакой передачи. Астроциты обеспечивают глютаматно-гамма-аминомаслянный кислотообмен. Поскольку глютамат и гаммааминомасл к-та похожи по структуре, и есть один фермент, кот делает из Глютамата ГАМК. Тут ещё и амины есть. И глиальные кл-ки имеют транспортные с-мы для перекачки медиаторов. Из них мож даже выделиться медиаторным способом экзоцитоза, как и у нейронов (на рис не показано).
Единственное отличие – астроциты не генирир-ют ПД!! Не генирируют спайков, поскольку между собой связаны стомозами – это фактически поры. Их (астроциты) можно фактически считать электрическими синапсами. Астроциты обеспеч важный процесс – кальциевые волны! А кальций – это непосредственный участник много чего в клетках. Во внеклеточ пространстве он имеет медиаторную природу, хотя сам ион не является сложным вещ-вом. Но тем не менее, к кальцию есть метаботропные рецепторы. И кальций здесь выступает как нейротрансмиттер (в широком понимании слова). Астроциты обесп некую цикличность кальциевых событий, они выбрасывают кальций в окр среду. Семьянов работает с глией много. Он говорит, что пора бы включить в вопрос пластичности астроциты, поскольку без них никакой пластичности быть не мож (может это оч категорическое заявление). Зелёный шар в верхнем правом углу – это пресинаптическое окончание. Тут как бы срезана с этой «пуговки» мембранка и показана везикула, наполненная различ-ми вещ-вами.
Астроциты необх для связи нейронов с кровенос руслом. Т.к. нейроны напрямую нигде не контагтир, кроме одного места – в гипоталамусе. И через астроцит происх весь транспорт того, что необх-мо нейрону.
Как генерируются ПД (спайки)?
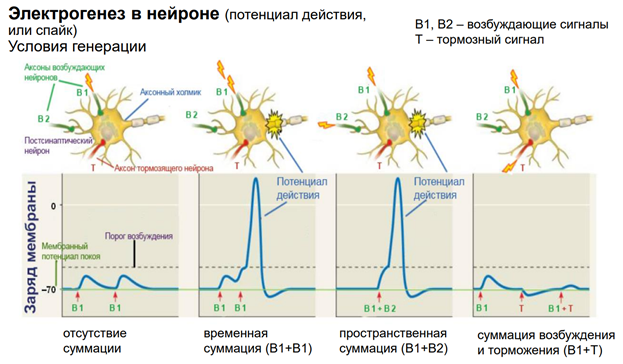
ПД обеспечивается потенциал зависимой проводимостью. Чтобы открылись Натриевые каналы, натриевый ток потек через клетки, необх некое возбуждение. Мы мож, конечно, электрическим способом раздражать нейрон, но в реальных нервных сетях нейрон получ некое синаптическое воздействие через возбуждающ или тормозные синапсы. См первую картинку слева: тут наверху изображен нейрон, у него есть аксонный холмик, где возникает ПД; два возбуждающих синапса (В1 и В2), кот мог расп-ся на дендритах/ соме. Если обеспечим раздражение через один вход подряд 2 раза, то эти два ВПСП обеспечат (если между ними достаточ длительн промежуток времени, поскольку ВПСП затухает из-за того, что нейромедиатор удаляется из синаптич щели – на это требуется какое-то время) не приведут к генерации ПД, т.к. потенциал на мембране не достигнет порогового. (См 2ую картинку слева.) Но если входы (В1 и В2) будем активировать через небольшое время, происх веменная суммация эффектов, достигнем порогов и получим ПД. Другой сп-б (третья картинка слева), когда мы активируем через два входа одновременно, тогда через мембрану потекут ионы одновременно (как бы в удвоенном кол-ве), достигнем порога, получим ПД. Пространственная суммация.
4 картинка. Если активируем вход, а затем тормозный вход, то быстро приведем к угнетению состояния нейрона, из-за того, что его мембранный потенциал сместится в отрицательн область относительно ПП (потенциала покоя), кл гиперполяризуется. И если мы одновременно будем раздражать возбудит и тормоз входы, не получим ВПСП. Такие процессы обеспеч перестройки активности нейронов.
Тут примерно о том же самом.
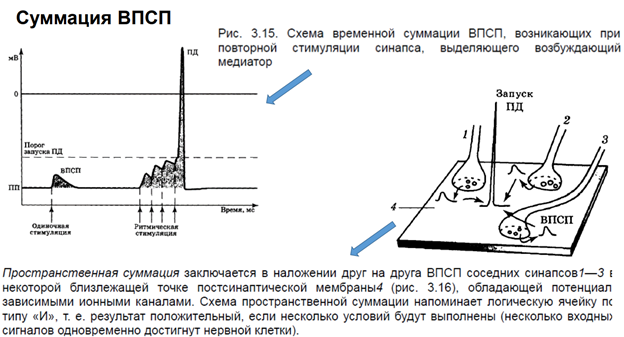
Это пример из ранних работ. На реальном объекте было получено, что одиноч стимуляция приводит к единичному ВПСП. А ритмическая стимуляция (= тетанус) приводит к потенциации синапса. Обычно ритмич стимул оказ большое вл на мембрану нейронов, поскольку обеспеч включение большого кол-ва синапсов. На левом рис изображен одиночный нейрон (где ритмическая стимуляция) беспозвоночного. Если дать несколько стимулов с высокой частотой, то ВПСП будут суммироваться, и в итоге обеспечат генерацию ПД, поскольку мембранный потенциал превысит порог. Это временная суммация. Пространственная суммация. Если все то же самое осуществим через разные синапсы. Активируем возбудительные, допустим, 1 и 2 синапсы одновременно или последовательно, но с малой частотой. Тогда каждый даст ВПСП. Если одновременно активируем, ВПСП будет большой. Есть трактовка через Бу’леву алгебру (ударение на У). Такая трактовка представляет каждую клетку (нейрон), как отдельную ячейку. И мы увидем результат только если будет активация нескольких клеток.
Посттетаническая депрессия
Кроме потенциации есть еще феномен постсинаптической депрессии. Это было сделано Эвартсом. Потенциация – это процесс в одну сторону, а депрессия – в др. Тут показан постсинаптический ответ латерального коленчатого тела после тетанизации контрлатерального оптического нерва у кошки в течение 20 сек. Исходные ответы были приняты за единицу. В течение длительного времени (почти 3 ч) наблюд депрессия потенциалов.
Феномен долговрем потенциации получаетчя прекрасно на разл участках коры ГМ (головного мозга), подкорков структурах. Это свид-во о том, что это универсаль мех-м, кот свойственен для многих синапсов.
Привыкание
Привыкание поведенческих ответов. Клеточн аналог привыкания
Костистые рыбы, включая Данио рерио, демонстрируют простую двигательную реакцию на испуг, которая контролируется двусторонней парой больших командных нейронов, клеток Маутнера, в заднем мозге рыбы. Двигательная реакция (C-start) запускается резким сенсорным (слуховым, визуальным или тактильным) стимулом. Эта реакция была зарегистрирована у личинок Данио на слуховой стимул через 4 дня после оплодотворения. Причём также было выявлено привыкание к стимулу. Функция данного поведение – избегание хищников.
Были описаны три формы привыкания: 1) Быстрое привыкание. (рис Б) Частые повторы стимулов – 50-120 шт за 1 мс (200Гц). После завершения тренировки через паузу в 3-15 мин поведенческий ответ возвращается к тому, котор был до привыкания.
2) Кратковременное. (рис А) Эффект сохраняется до 1 ч после тренировки. Предъявление стимулов блоками (всего их 10) по 900 шт (1Гц), интервал между блоками 5 мин. Было обнаружено, что для данного привыкания необходима активация N-метил-d-аспартата (NMDAR). 3) Длительное привыкание. Стимулировали 6 блоками, разнесённые во времени; в каждом блоке по 8 коротких слуховых стимулов (1 Гц). Привыкание сохранялось до 18 ч. Индукция данного привыкания также зав-т от NMDAR.
Что по рисунку Б: возврат к предтренировочному уровню через 15 мин. Звездочка указывает, что 10-секундный тест значительно отличался от предварительного теста и 15-минутного теста. Решетка – указывает, что 1-минутный тест от предварительного теста и 15-минутного теста.
* Сенсабилизация – форма неассоциатривного обучения, при которой воздействиее возбуждающего стимула вызыв усиление реакции. Если мы этим же стимулом усиливаем привычную реакцию – это дезабитуация ( DISHABITUATION).
Интересно, что введение DL-2-амино-5-фосфонопентановой кислоты вместе с никотином блокировало сенсибилизацию. Следовательно, активность NMDAR, видимо, опосредует некоторые формы привыкания и чувствительности.
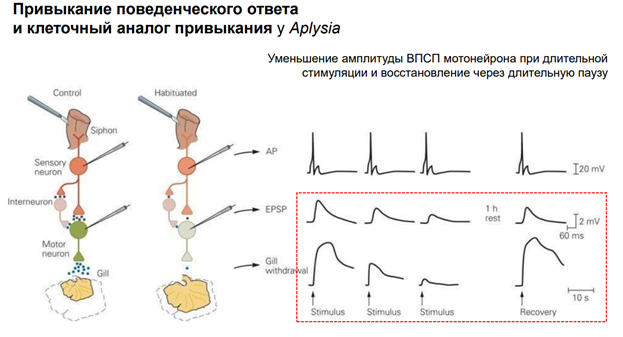
Объект: морская улитка Aplysia californica. Привыкание - это снижение реакции на повторное применение данного стимула, которое нельзя отнести к сенсорной адаптации, сенсорной или моторной усталости (Thompson and Spencer, 1966; Rankin et al., 2009). Интенсивность стимулов (последовательности 500 мсек [25 Гц] импульсов тока 10 мсек) устанавливалась для каждого препарата выше порогового значения для надежного вызова отдергивания жабр и оставалась неизменной на протяжении всего эксперимента. Схема эксперимента: Стимулировали сифон – две стороны: контрольная и стимулируемая (=тренированная). Предтренировка: Стимул в тренируемую сторону, через 5 мин в другую (контрольную) сторону. Ждали 1 час, потом эксперимент. Всего было 5 блоков стимуляции. Интервал межблочный = 90 мин. Во время каждого блока тренируемой стороне давали 30 стимулов. Далее после 2 ч отдыха обоим сторонам давали по 1 стимулу (с разницей во времени 5 мин). Результаты: Тренировка привела к долгосрочному привыканию рефлекса отвода жабр к стимуляции тренированной стороны сифона. Закрашенные кружки представляют реакцию на стимуляцию тренированной стороны, тогда как закрашенные квадраты представляют реакцию на стимуляцию нетренированной стороны.
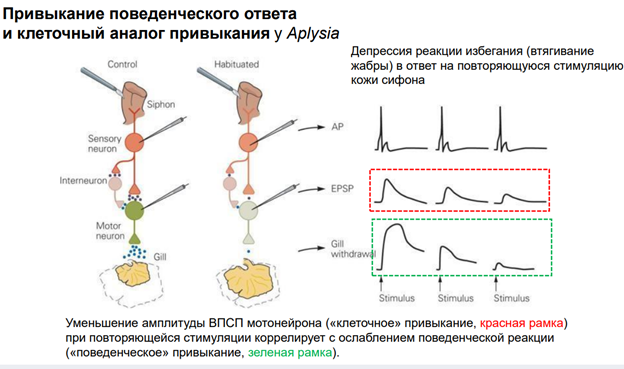
Механосенсорные нейроны запускают высокочастотные импульсы в ответ даже на умеренные тактильные стимулы на кожу. Сенсорные нейроны также запускают вспышки в диапазоне 1-60 Гц в ответ на электрич стимулы. Метод. Электроды были 1 в сенсорном нейроне, 2 других (регистрирующий и вводящий ток) мотонейронах. Двигательный нейрон был гиперполяризован в теч всего эксперимента до -80мВ, чтобы предотвратить суммацию ВПСП от запуска ПД. Сенсорный нейроне стимулировали 1 сек при частоте 10 Гц (10 мс импульсы, 100 мс межимпульсн интервал) током, котор в 1,3 раза превышал пороговый. Далее между блоками использовался интервал в 10 мин, чтобы синапс мог восстановиться. Всего было проведено 10 стимуляций. Результат: Сенсорные нейроны запускают всплески ПД при активации механическими стимулами, приложенными к коже. Рис 1А: ответ сенсорного нейрона сифона на серию контролируемых механич стимулов на коже. Шаг между воздействиями 800 мс, кле точный разряд увеличивается в зависимости от интенсивности стимула. Рис. Б: хвост стимулируют, полуинтактный препарат. 200 сек стимуляция длится (ее начало обозначено стрелкой - shock). Сенсорный нейрон срабатывает с частотой, соответствующей стимулу (60 Гц), с небольшой адаптацией. За этим разрядом следует другой разряд, во время которого частота сот-ет 10 Гц.
На рис А: пример всплеска ПД, вызванного в плевральном сенсорном нейроне (SN) и результирующего суммиранного ВПСП в двигательном нейроне (MN). Амплитуды отдельных ВПСП снижаются до установившегося уровня 20% от начального ВПСП. Постсинаптический ответ был разделен на три фазы: инициация ВПСП, переходная фаза (со 2 по 5 зубец) (в теч которого синаптич эффективность снижалась) и фаза устойчивого состояния (с 6 по 10 ВПСП) (в теч которого амплитуды ВПСП достигли стабильного уровня депресси). Рис Б: совокупные данные для всплесков 10 Гц, в котор амплитуды ВПСП были нормализованы к амплитуде начального ВПСП.
В эксперименте проанализировали 40 клеток (14 ЛПа3 и 26 ППа3). Мембранный потенциал кл-ок варьировался от -45 до -85 мВ.
NCR – никотиновые рецепторы
MCR – мускариновые рецепторы
Анатомическая специфика. В экспериментах были идентифицированы эти синапсы. Они оказались холинэргическими. Были идентиф-ны холинэргические нейроны. И, соотв-но, рецепторы мембран этих нейронов. И для имитации таких возд-ий был использован искусственный синапс. К тому месту, где на мембране нейрона раса-сь АЦХые рецепторы подводилась пипетка, наполненная АЦХ. И методом микроионофореза мож было выделять из пипетки дозированное кол-во АЦХ на пов-ть нейронов. 1) Можно было менять концентрацией АЦХ в пипетке. 2) Мож было варьировать длительностью и интенсивностью тока для электрофореза. Чем больше ток, тем больше Ацх выделится. Т.о. иск синапс заменял реальный. На нейрон иммитировали тактильное воздействие кол-вом ацх, которым воздействовали. При этом были зарег-ны токи (методом фиксации потенциала). Надо было исп пипетки тонкие. Метод пэч-клямп целой клетки был, можно было фиксир потенциал на нуж уровне и рег-ть токи
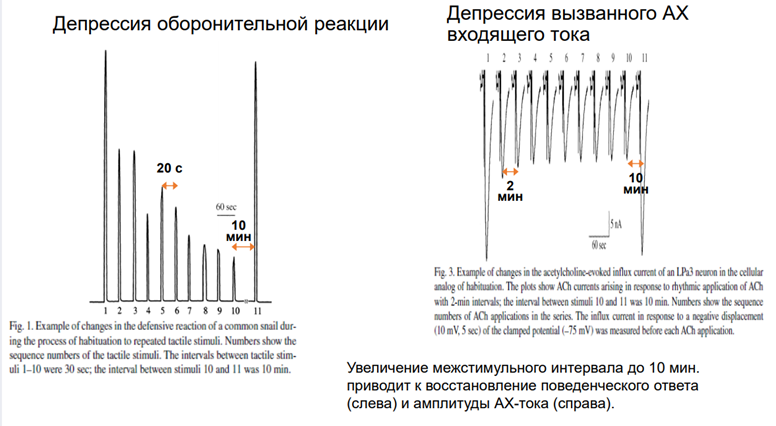
Картинка справа. Регистрировали токи. Интервал между воздействиями ацх-ом 2 мин. Токи входящего направления, т.к ацх рецептор обеспеч ВПСП, токи катионовые д.б входящими. На первую аппликацию ацх-ом получаем высокоА ток. Если через 2 мин повторяем стимул-ю, ток уже меньше. На 4 и 5 наступает плато. Если сделаем перерыв, получим восстановление А. Заключение: таким поведением управляет командные нейроны, актив-ся ацх-ом.
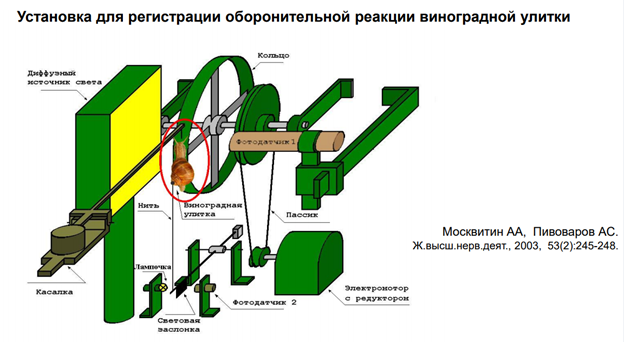
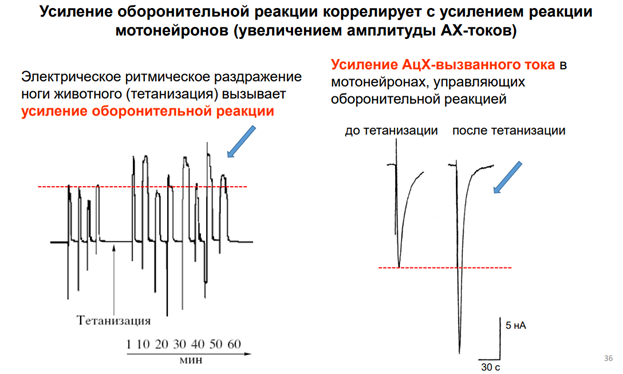
Чечь идет о сенситизации оборонитльной р-ии. При касании улитки за антенулы, она сожмется, но не до конца. А если проведём тетанизацию по хвосту, и после неё осущ-м тактильную стимул-ю, увидим р-ю большей А.
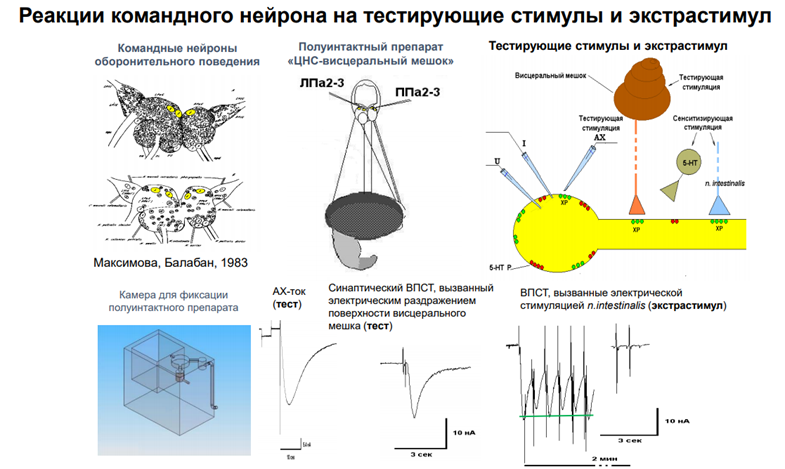
Тут есть картинка из работы Балабана. Командные нейроны были помечены. Сверху фото ганглиев (серые такие), а снизу схема. И тут показано, где они нах-ся (эти нейроны). Под ними (синяя картинка) – установка. Левее – АЦХ токи, которые былм зарег-ны. Синаптич ток, вызванный электрич раздражением пов-ти висцерального мешка (тест). Теперь смотрим в верхний правый угол. Тут висцеральный мешок. Который надо простимулировать, тогда активир-ся аксоны (оранжевый пунктир), котор выделяют ацх, и на дендритах командного нейрона обеспечивают такие ответы. Есть еще стимуляция через др нерв (голубой). Он является экстрастимулом, т.к по нему потенциально мож прийти сеситизирующая стимуляция. И она мож вызввать тоже какой-то ацх-ый ответ. А пунктиром он нарисован, потому что это высказано науровен предположения, т.к в работах Пивоварова эти пути не были прокрашены!. И считается, что стимуляция по этому пути не вызыв АЦХ токов, хотя это холинэргические синапсы. Итак, у нас есть тестирующая стимуляция из висцерального мешка (воздействие касалки). А еще есть искуственный ацх синапс. И везде рецепторы одни и те же, НО есть данные, что ацх, кот действует на местрану нейрона может быть не синаптическим. Он мож выделяться в среду из мешка (без синапса). Т.е. в данном сл медиатор выступает как гормон, и не обязательно он долджен действаовть через синапс. У моллюсков это норма. У них мож быть как синапс, так и вот такое воздействие. На этом рис-ке еще есть серотонин, и на этом нейроне есть серотониновые рецепторы (красные), но не идентифицирован контакт от серотониноваого нейрона. Т.е он может выступать как гормон – выделяться просто в межклеточную жидкость и действ на мембрану.
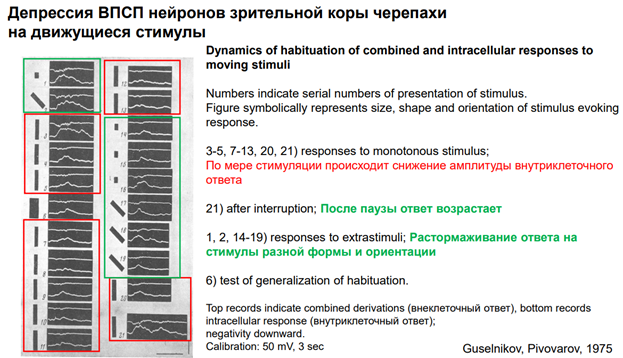
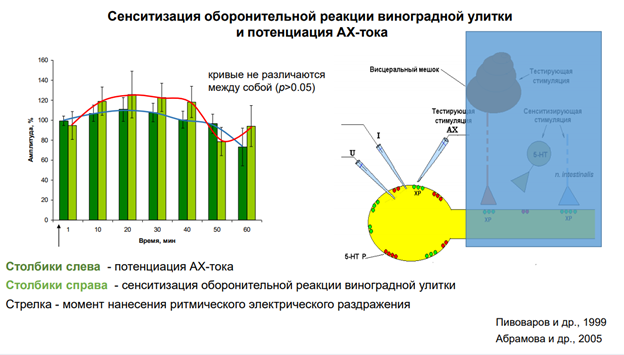
Если сопоставить исследования, проведенные на поведенческом и на нейронном уровне. Тут показана сенситизация оборонительной р-ии улитки. Использовалм 10-12 животных. Оу -относительная шкала, А этой р-ии. Зеленые столбики – опытная серия. Через 10 мин давали тестовый стимул, а до этого проводили сенситизацию (вертикальная черная стрелка вверх). Видим, что на 10-20-30 мин получается хорошая сенситизация. К 50-60 минуте сенситизация исчезает. Желтые столбикик – контрль – через каждые 10 мин дается стимуляция, наблюдаем депрессию ответа, привыкание.
Правее изображены единичные записи конкретного эивотного. Вниз пики резкие – это артефакты от пишущего пера чернильного (ну а что вы хотели 😊, старая работа). Они получены на самописце. Это ток, котор течет, генерируется фотодатчиком, отражает уменьшение площади тени молюска (чем меньше моллюск, тем сильнее р-я). Зеленые – контроль, потом тетанизация, и то, что под оранжевой чертой – это процес сенситизации – в течение длительного времени животное демонстирует такую динамику. Полосы эти горизонтальные проведены ЛАтановым на глазок. Если это просуммировать данные по 10, напр, животным, то получим что в контрле меньше действительно А. А если посмотреть контроль, когда нет никакой сенситизации (картинка правее, где только зеленая черта), получаем что общ уровень р-ии один.
Что с токами получается? Справа картинка. До тетанизации ток был небольшой, а после в 2 раза по пиковой А увеличился
Если сопоставить данные поведения и клеток. Р-я не очень выражена. Выпуклость оранжевая не оч яркая, но если провести корелляционный анализ, то кривые эти не различаются. Это примерно одно и то же. Может это не особо корректно сравниватть… Но сравнили. И получили, что они дстаточно скореллированы. Это очень хороший клеточный аналог сенситизации. Сенситизация – вариант неассоц научения или неассоц памяти
Классическое обуславливание
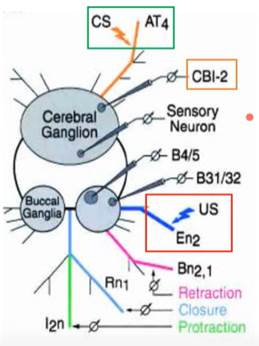
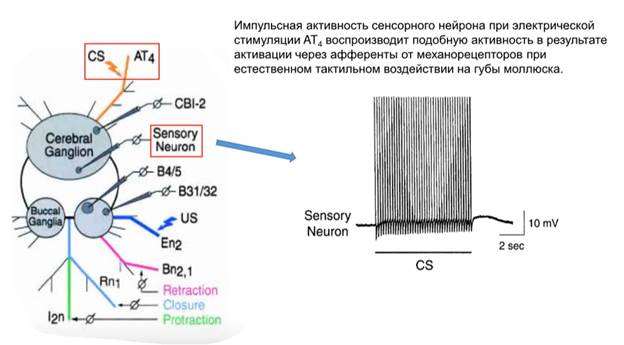
Рассматривается пищевое поведение аплизии, при этом разделяют воспринимающую часть этого поведения и эффекторную. В качестве УС (CS, condition stimulus) используют электрическое раздражение губного нерва (АТ4). В качестве БС (US) – стимуляция En2 (нерва, участвующего в реализации пищевого поведения, называется esophageal nerve). Был найден командный нейрон, CBI-2, он активировался при стимуляции AT4 (УС).
** Retraction, closure, protraction – реакции, входящие в состав т.н. buccal motor pattern – пищедобывательного паттерна, запускающегося через буккальный ганглий при активации нерва En 2. Переводятся как растягивание, закрывание и втягивание [радулы и одонтофора].
1 – УС и БС длятся 8 и 4 сек, с перекрытием (сочетание) 2 - «проверка на псевдообуславливание», стимулы той же длительности предъявляют по одиночке, м.б. даже в случайном порядке 3 – предъявляют только БС
В 1 случае мы ожидаем условную реакцию, в 3-м – безусловную.
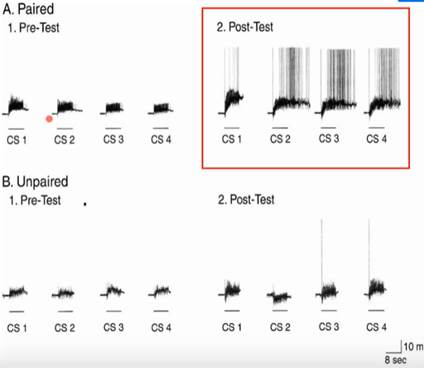
Эксперимент: CSs – просто несколько предъявлений УС; в одной сессии после претеста идут предъявления (training) – 10 сочетаний УС+БС с интервалами 4 мин между ними, потом моллюск час думает о своём, и после пост-теста сессия заканчивается.
И что получилось? Во-первых, оказалось, что при стимуляции АТ4 сенсорный нейрон реагирует так же, как если бы что-то реально потрогало губу моллюска:
Но нам ведь интересен в первую очередь командный нейрон:
P – протокол paired, UP – протокол unpaired (т.е. было 12 регистраций первого и 13 второго); UP в данном случае – контроль, параметры работы нейрона в норме (помним, он возбуждается при УС, про него и речь). Resting membrane potential – ПП, input resistance – входное сопротивление; за 100% принимали параметры, измеренные до начала эксперимента. Изменения оказались в пределах ошибки.
А что по испульсной активности?
После сочетаний активность нейронов CBI-2 значительно усиливается, эффект наблюдали на 6/12 нейронах. В протоколе Unpaired такого эффекта не было ни у одного нейрона. Ну и на уровне пищевых реакций тоже было, на что посмотреть:
Звёздочками (которые выглядят, как кружочки) отмечены те реакции, которые есть в пост-тесте по сравнению с пре-тестом. Протоколы B и C нам мало интересны, важнее, что при сочетании УС с БС есть отчётливая разница в реакциях на УС между теми, что были до и после сочетаний. Оранжевое – артефакты от стимуляции АТ4.
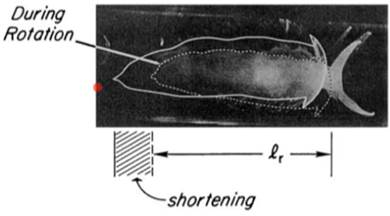
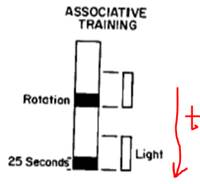
В ответ на вращение моллюск демонстрирует конкретную реакцию, смысл которой – уменьшить площадь своего тела, когда волны создают турбулентность. А именно – он укорачивает ногу и цепляется ею за субстрат. Вращают бедолагу на скорости 200 об / мин.
Свет является для него индифферентным стимулом (ИС), но в ответ на него нога удлиняется (зверушка проявляет фототаксис). Вырабатываем УР
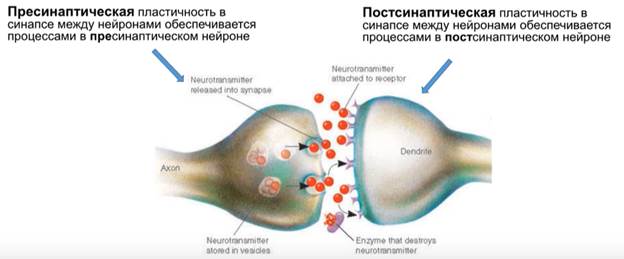
Синаптическая пластичность
Системная пластичность – понятие сложное и выходящее за рамки ЦНС, она включает гомеостаз. Про неё не говорим Говорим о пре- и постсинаптичесой пластичности между нейронами на клеточных аналогах научения.
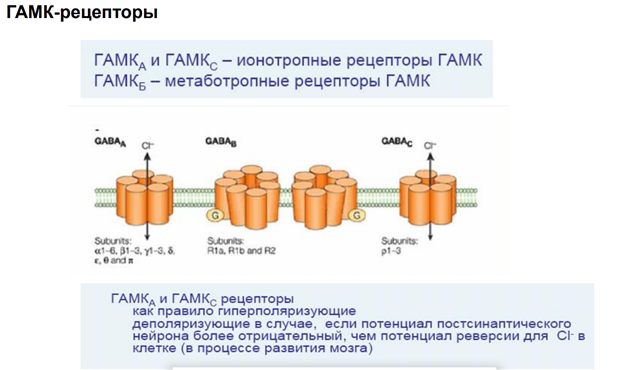
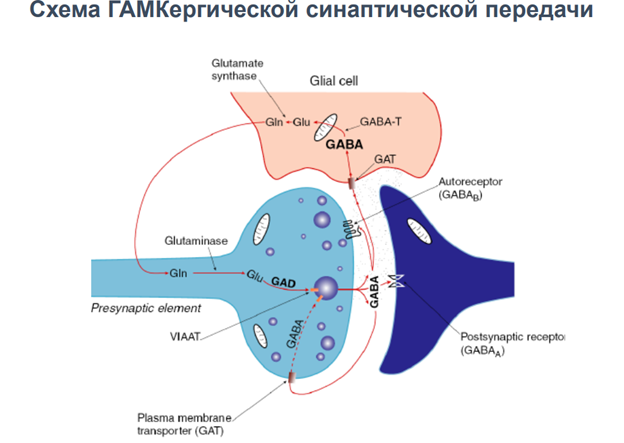
ГАМК-рецепторы
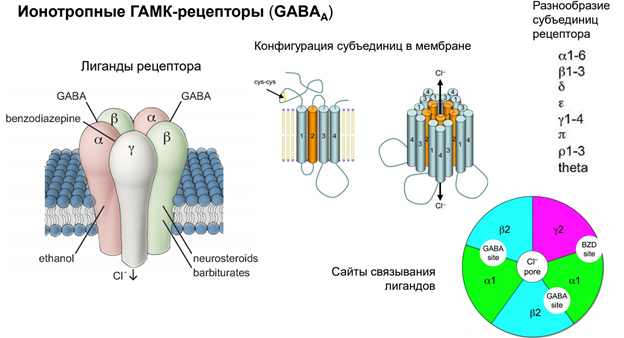
Гамк рецептор. В его состав 5 субъединиц. Каждая представлена 4 транс мембранными участками. Много субъединиц. Варианты его разнообразны поэтому. Есть места связывария для барбитуратов (бензодезепиновая субъединица, выполняет тебя же эффекты что и гамк, наличие бензодиазепинов усиливает эффект, выполняет ф-ю ко медиаторов). Блокатор - микро токсин (прекращает проводимость через холодный канал).
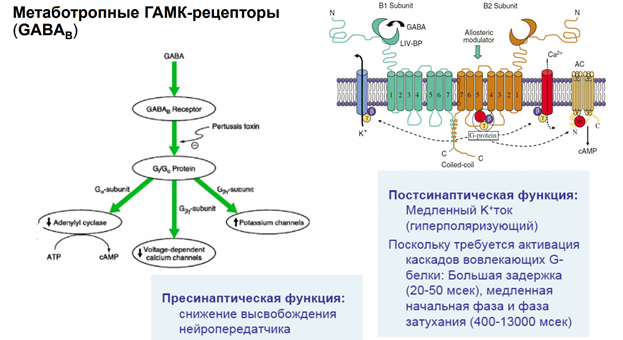
На всех синапсах есть рецепторы для связывания ацх. Эти метаботропн ведь рецепторы, который активирся гамком. Ретроградная регуляция выделения нейромедиатора. Обеспечивает снижение выделения самой себя. Блокируется кальциевый канал, который зап экзоцитоз. Вероятность уменьшается выхода кальция.
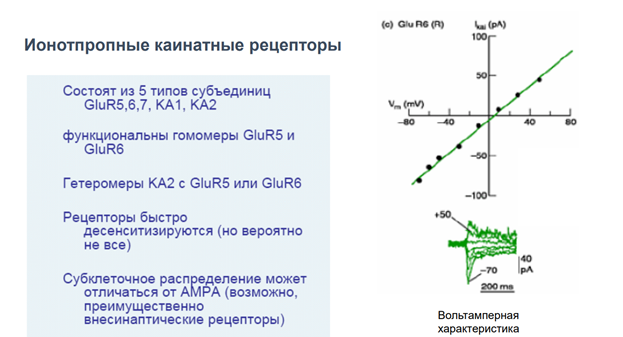
Глутаматные рецепторы
Ионотропные АМПА-рецепторы
Метаботропные рецепторы
Одни из Глютаматных рецепторов метаботропных обеспечивает пластические перестройки на клетках Пуркинье мозжечка, обеспечивая синаптическую дипрессию.
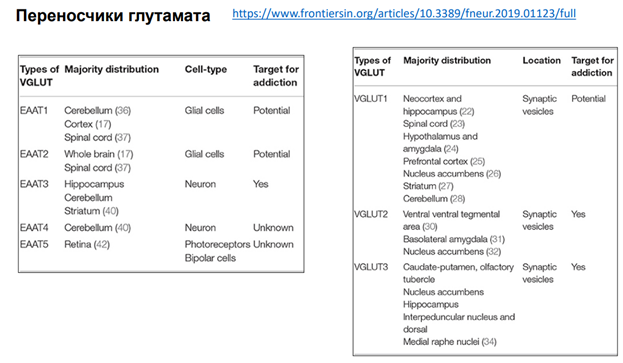
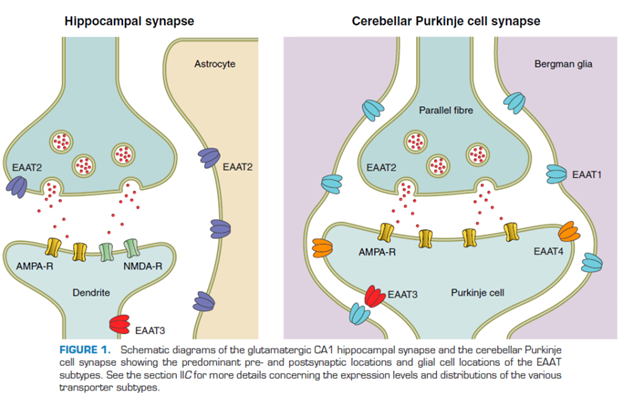
https://www.frontiersin.org/articles/10.3389/fneur.2019.01123/full Кроме ионных процессов, активации рецепторов очень важны другие участники синаптич передачи – переносчики: глютамата, нейромедиаторов. Они относятся к ионным обменникам, ко вторичной системе переноса (первичные процессы с участием АТФ происходят, т.к для работы этих насосов (попм) требуется молекула АТФ и первичн системы мембранного транспорта – обеспечивают ионный градиент, необх-ый для функционирования клеток; а вот уже ионные градиенты используются другими переносчиками, которые используют энергию от разности концентраций). Переносчиков огромное кол-во. В левой таблице переносчики Глутамата (через мембрану), а в правой – везикулярные переносчики (перетаскивают Глютамат в синаптические везикулы). Переносчики Глутамата через мембарну (5 шт в таблице) локализованы: 1 и 2 тип в глиальных клетках локализованы, 3 и 4 – в нейронах, 5 – в фоторецепторах сетчатки. Везикулярные переносчики – были обнаружены в синаптических везикулах.
Тут показано, где локализованы эти переносчики. А астроцитах, … (см слайд выше). Астроциты извлекают из синаптич щели глютомат. Вовлекают в его в цикл, чтобы потом вернуть обратно. В клетке Пуркинье тоже хорошо представлены (картинка слева). Бергманская глия – в принципе, выполняют такую же функцию, как и астроциты, просто у них другая локализация и есть некоторые особенности.
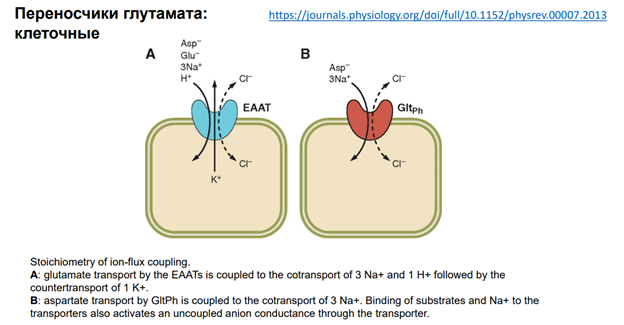
https://journals.physiology.org/doi/full/10.1152/physrev.00007.2013 Переносчики работают за счет градиента Калия и иногда за счет градиента Хлора, а также тут может участвовать Натрий, как главная движущая сила.
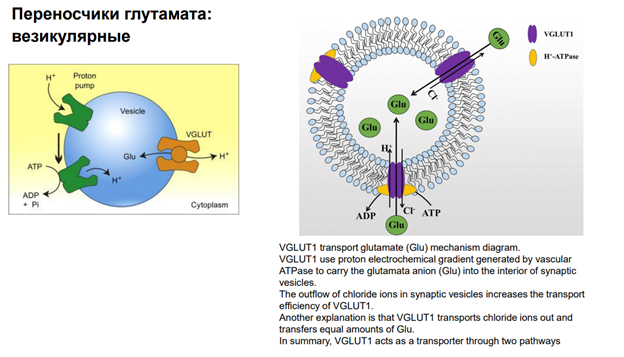
Смотрим левую картинку: Перед тем, как переносить Глютамат, надо создать протонный градиент, поскольку везикулярные переносчики используют градиент протонов! (а не градиент каких-то ионов) Чтобы создать это градиент необходима система первичного транспорта. И вот есть H- ATP-аза (Эйч-АТээФаза), которая закачивает в везикулы протоны. И благодаря выходу протонов (они выходят по своим концентрационным градиентам), глютамат попадает в везикулу. На правой картинке аналогичная схема.
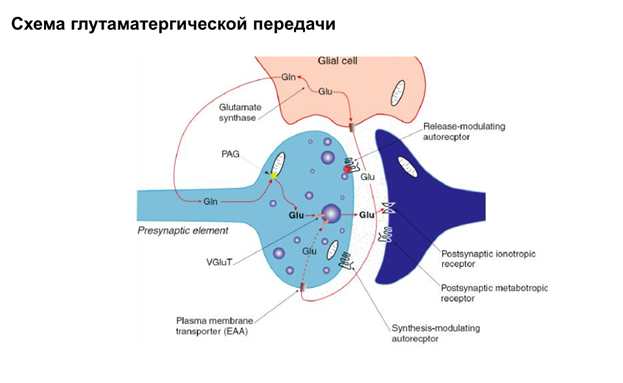
Глтаматный синапс. Глутамат переносчиком возвращается в глиальную клетку. Он может вернуться также и в саму пресинаптическую клетку. Иногда он может попасть и в постсинаптическую клетку.
|
|||||||||
|
|
Последнее изменение этой страницы: 2022-01-22; просмотров: 28; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.140.186.201 (0.228 с.) |




























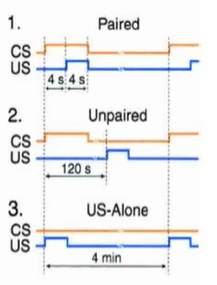 Протоколы подачи стимулов:
Протоколы подачи стимулов: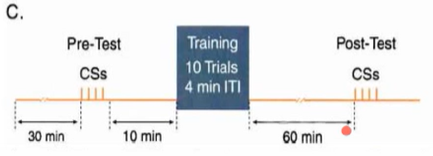








 У моллюска есть глаза, статоциты (аналог вестибулярного аппарата), и ЦНС с 2-мя крупными ганглиями.
У моллюска есть глаза, статоциты (аналог вестибулярного аппарата), и ЦНС с 2-мя крупными ганглиями.