Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
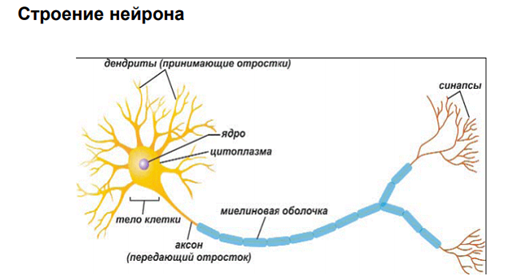
Пластичность нервной системы
Пластичность нейрона. В резул перестроек нейрон может что-то усилить (потенциация) или ослабить (депрессия).
У нервных кл-к есть ещё такие функциональные характеристики, как перекачивание ионов против концентрационного градиента, что составляет возможность существования этих клеток. Нерв клетки пропускают по своим каналам ионы в обе стороны – основная и наиб специф-ая хар-ка. Конорски и Костюк. Они определяют пластичность как длительн модификации клеток и синаптич функции. Они говорят, что в основе модификаций поведения лежат перестройки в нейронных сетях. Основные феномены, кот отраж-ют изменение эффективности синапт передачи: · Длительн потенциация · Гетеросинаптич облегчение · Неассоциатив формы научения. В общей классификации памяти их называют стимулзависимое поведение. Оно не ассоциативное, т.к нам достаточно всего одного стимула, чтобы получ на него привыкание/ сенситизацию. Для сенситизации нуж опред условия (далее) · Ассоциатив научение, котор представлено рефлексами различной сложности – классический и инструментальный.
Эрик Кэндэл. Нобль лауреат. Обобщил многолетний опыт своих предшественников. С 60х работал в области синапт пластичности. Аплизия – большой моллюск. На нем изучали не ассоциативные формы научения. Написал книгу «Клеточные основы поведения», тут значит часть посвящ пластичности. Дэвид Гланзмэн. Работал
Наши соотечественники, котор внесли опред вклад в исследование пластичности. Соколов. Латанов работал с ним более 10 лет в области цветового зрения. Заложил основы в представлении... Он не занимался пластичностью напрямую, но он дополнил современными даннымимодель рефлекторной дуги (дополнил Павлова). Он был заведующим кафедры Психофизиологии МГУ. Сахаров. Занимался беспами. Написал книгу «Генеология нейрона», где он описывает эволюцию развития мех-мов, кот опред пластические перестройки беспов. Есть значимые мания, как нейрон интегрирующ информацию, кот в него поступает. Работал на простом поведении моллюска (это хорошо, т.к. простые Нервные с-мы (НС) дают более однозначный резул-т, хотя более редукционистский). Он считал, что все дело в медиаторах. И пластичность опред-ся различными воздействиями со стороны разл медиоторных с-м. И вступал в полемику с теми, кто рассм ПД, как отражающие вопросы пластичности. Но и без Сахорова понятно, что ПД (потенциалы действия) – это способ распространения инфо на (относительно) далёкие расстояния. И сам ПД – все или ничего. И если «все или ничего», то у него два состояния – «ноль» и «единица». И как мож быть закодирована инфо, кот поступает в ПД? Никак! Может, если говорить о числе спайк ов, их продолжительности, паче и т.д. Но все, на сам деле, просих на мембранах нейронных кл-к, на синапсах. И синаптич паттерн активации и явл тем феноменом, кот отраж содержательные некие имения в мембране нейронов.
Балабан. Пивоваров. Работал на отдельных клетках
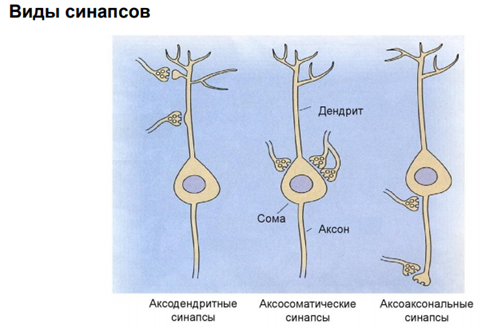
На сам деле такие сп-бы контактов – это такой сп-б разделить потоки инфо-ции, которая так или иначе влияет на состояние нейрона, его функционирование в сетях нейронов. Синапсы быв разные по их локализации, эргичности, морфологии…
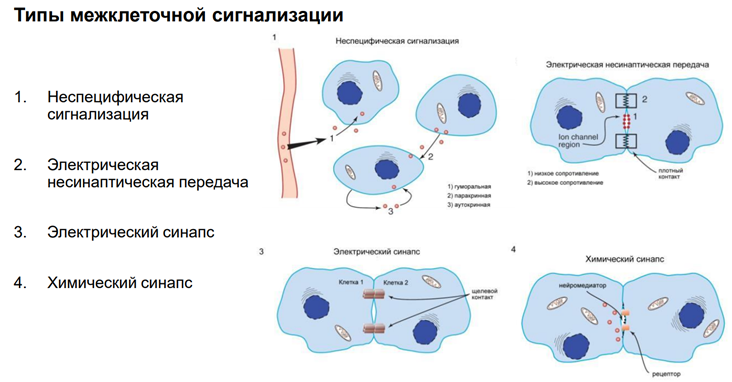
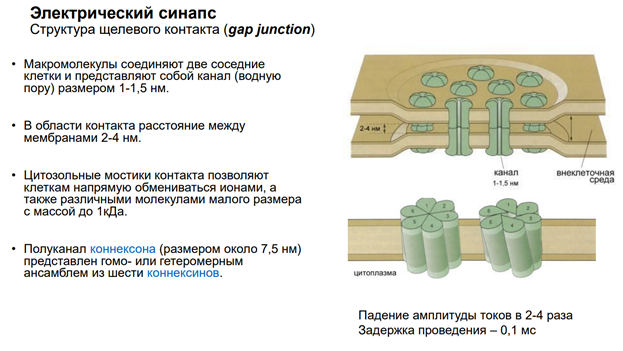
Мы в основном будем говорить про химический синапс. Но есть ещё неспецифич сигнализация. Когда агент, который влияет на нейрон, им происхождение не из аксонов, а из крови, неклеточной жидкости. Это гуморальная (паракринная и аутокринная) регуляция. Так действуют гормоны. Здесь неспецифическая связь. Эффект определяется только тем, есть ли рецептор на клетке к этому вещ-ву или нет. Электрич синапсы. Нам в этом курсе не интересны, т.к. синаптич перестроек тут не следует ожидать из-за того, что тут простые образования – представляют из себя «провода», котор соед-ют клетки. А химический синапс нам интересен, т.к. многообразие медиаторов / веществ, которые имеют нейромедиаторную природу, обеспеч разнообразные эффекты
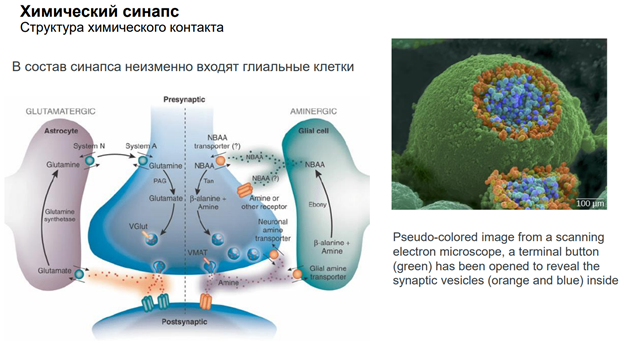
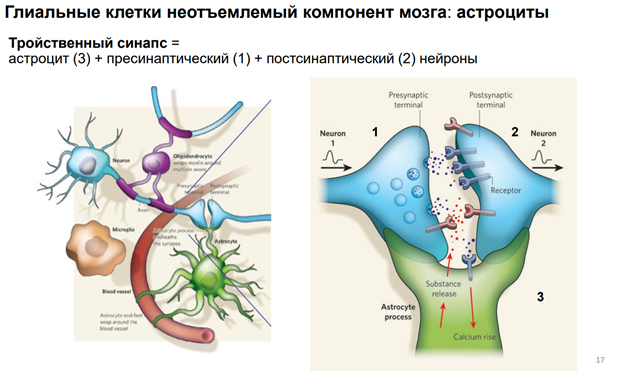
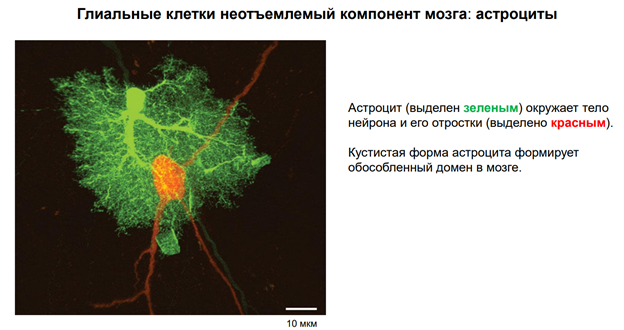
Глия. Сейчас к ней много внимания. В частности, к астроцитам, кот являются участниками синаптич передачи. На картинке показано, как астроциты окружают клетку. Астроциты бывают глутаматэргические и аминэргические. Они происх из общего предка в эмбриогенезе (из стволовой кл). Они ближайшие родственники нейрона. Астроциты имеют фактически все св-ва нейронов. Там есть и транспорт с-ма. Например, без астроцита не может осущ-ся глютаматэргическая передача, поскольку предшественники глутамата синтезируются в специальном астроците (розовый слева на картинке), и без него не могло бы быть никакой передачи.
Астроциты обеспечивают глютаматно-гамма-аминомаслянный кислотообмен. Поскольку глютамат и гаммааминомасл к-та похожи по структуре, и есть один фермент, кот делает из Глютамата ГАМК. Тут ещё и амины есть. И глиальные кл-ки имеют транспортные с-мы для перекачки медиаторов. Из них мож даже выделиться медиаторным способом экзоцитоза, как и у нейронов (на рис не показано). Единственное отличие – астроциты не генирир-ют ПД!! Не генирируют спайков, поскольку между собой связаны стомозами – это фактически поры. Их (астроциты) можно фактически считать электрическими синапсами. Астроциты обеспеч важный процесс – кальциевые волны! А кальций – это непосредственный участник много чего в клетках. Во внеклеточ пространстве он имеет медиаторную природу, хотя сам ион не является сложным вещ-вом. Но тем не менее, к кальцию есть метаботропные рецепторы. И кальций здесь выступает как нейротрансмиттер (в широком понимании слова). Астроциты обесп некую цикличность кальциевых событий, они выбрасывают кальций в окр среду. Семьянов работает с глией много. Он говорит, что пора бы включить в вопрос пластичности астроциты, поскольку без них никакой пластичности быть не мож (может это оч категорическое заявление). Зелёный шар в верхнем правом углу – это пресинаптическое окончание. Тут как бы срезана с этой «пуговки» мембранка и показана везикула, наполненная различ-ми вещ-вами.
Астроциты необх для связи нейронов с кровенос руслом. Т.к. нейроны напрямую нигде не контагтир, кроме одного места – в гипоталамусе. И через астроцит происх весь транспорт того, что необх-мо нейрону.
Как генерируются ПД (спайки)?
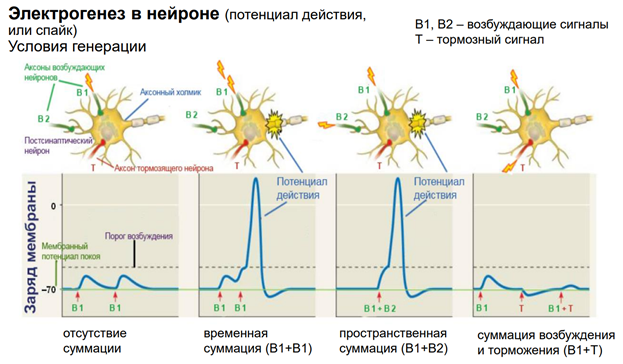
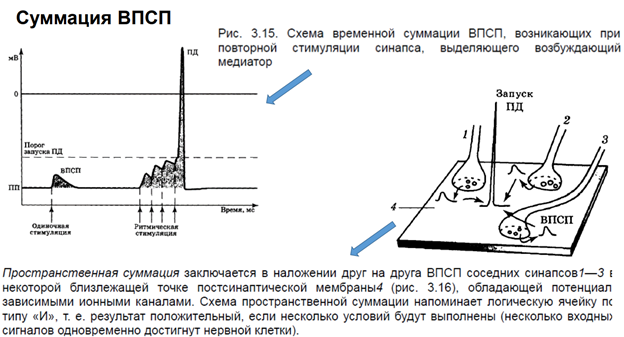
ПД обеспечивается потенциал зависимой проводимостью. Чтобы открылись Натриевые каналы, натриевый ток потек через клетки, необх некое возбуждение. Мы мож, конечно, электрическим способом раздражать нейрон, но в реальных нервных сетях нейрон получ некое синаптическое воздействие через возбуждающ или тормозные синапсы. См первую картинку слева: тут наверху изображен нейрон, у него есть аксонный холмик, где возникает ПД; два возбуждающих синапса (В1 и В2), кот мог расп-ся на дендритах/ соме. Если обеспечим раздражение через один вход подряд 2 раза, то эти два ВПСП обеспечат (если между ними достаточ длительн промежуток времени, поскольку ВПСП затухает из-за того, что нейромедиатор удаляется из синаптич щели – на это требуется какое-то время) не приведут к генерации ПД, т.к. потенциал на мембране не достигнет порогового. (См 2ую картинку слева.) Но если входы (В1 и В2) будем активировать через небольшое время, происх веменная суммация эффектов, достигнем порогов и получим ПД. Другой сп-б (третья картинка слева), когда мы активируем через два входа одновременно, тогда через мембрану потекут ионы одновременно (как бы в удвоенном кол-ве), достигнем порога, получим ПД. Пространственная суммация. 4 картинка. Если активируем вход, а затем тормозный вход, то быстро приведем к угнетению состояния нейрона, из-за того, что его мембранный потенциал сместится в отрицательн область относительно ПП (потенциала покоя), кл гиперполяризуется. И если мы одновременно будем раздражать возбудит и тормоз входы, не получим ВПСП.
Такие процессы обеспеч перестройки активности нейронов.
Тут примерно о том же самом.
Это пример из ранних работ. На реальном объекте было получено, что одиноч стимуляция приводит к единичному ВПСП. А ритмическая стимуляция (= тетанус) приводит к потенциации синапса. Обычно ритмич стимул оказ большое вл на мембрану нейронов, поскольку обеспеч включение большого кол-ва синапсов. На левом рис изображен одиночный нейрон (где ритмическая стимуляция) беспозвоночного. Если дать несколько стимулов с высокой частотой, то ВПСП будут суммироваться, и в итоге обеспечат генерацию ПД, поскольку мембранный потенциал превысит порог. Это временная суммация. Пространственная суммация. Если все то же самое осуществим через разные синапсы. Активируем возбудительные, допустим, 1 и 2 синапсы одновременно или последовательно, но с малой частотой. Тогда каждый даст ВПСП. Если одновременно активируем, ВПСП будет большой. Есть трактовка через Бу’леву алгебру (ударение на У). Такая трактовка представляет каждую клетку (нейрон), как отдельную ячейку. И мы увидем результат только если будет активация нескольких клеток.
|
|||||||
|
|
Последнее изменение этой страницы: 2022-01-22; просмотров: 34; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.149.251.154 (0.01 с.) |