Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
I. Теорія гістогенів Ганштейна, 1863 р.Стр 1 из 4Следующая ⇒
Гістогенами називаються спеціалізовані меристематичні зони. Отже, точка росту стебел складається з ініціальних клітин, що постійно діляться мітотично. Трохи нижче ініціальні клітини починають збільшуватись, диференціюватись. Вони поділяються мітотично, але закладається диференціація в напрямку постійних тканин (зона промеристеми). Нижче розташовується зона первинної меристеми, де виділяються клітини, які дають покривну тканину і центральний циліндр. В корені такими зонами є: 1. дерматоген – перший шар клітин; 2. периблема – яка дає первинну кору; 3. плерома – яка даватиме центральний циліндр і провідні елементи. II. Теорія «корпуса і туніки» Шмідта, 1924 р. Згідно з цією теорією, апікальна меристема складається з двох масивів меристематичних тканин або з двох зон – туніки і корпуса. Туніка розміщена ззовні – периферична меристема, і буває одно-, дво- і багатошаровою. Характерною її особливістю є те, що її клітини поділяються тільки в одному напрямі – уздовж поверхні апексу (тобто тангентально). Якщо туніка одношарова, то з даного шару утворюється епідерміс. Якщо багатошарова, то або дає первинну кору повністю, або частково. Під шаром туніки знаходиться корпус, клітини якої поділяються у всіх напрямках, забезпечуючи об’ємний ріст апексів. Корпус дає центральний циліндр. Якщо туніка одношарова, то корпус буде формувати і первинну кору або її частину. При утворенні епідерми кореня з дерматогена поділ клітин проходить антикленально (перпендикулярно радіусу) і перикленально (паралельно радіусу). В зоні промеристем в центральній частині виділяється зона сильно видовжених клітин, і якщо всі інші ізодіаметричні, то видовджені стають прозенхіматичними і називаються десмогеном. Десмогени в зоні первинної меристеми чи в зоні корпуса стають ще видовженішими, які в межах первинної меристеми називаються прокамбієм – зона переходу від апікальної до латеральної зони. ПОКРИВНІ ТКАНИНИ
На поверхні всіх органів рослин знаходяться покривні тканини. Основна їх функція захисна. Захищають внутрішні тканини від несприятливих факторів: механічних впливів, різкої зміни температури, проникнення шкідників та патогенних мікроорганізмів, зайвого випаровування і висихання. Крім захисної покривні тканини виконують ще й інші функції: газообміну, транспірації, виділення, поглинання.
За походженням покривної тканини розрізняють: 1. первинну – епідерма (або епідерміс), що походить від туніки або дерматогену; 2. вторинну – перидерма, що походить від фелогену (коркового камбію); 3. третинну – ритидон або кірка, що є ситемою перидерм.
Епідерма або шкірка – це первинна покривна тканина. Вкриває листки, квітки, плоди, а також стебло при їх первинній будові. В процесі еволюції епідерма виникла при виході рослин на сушу, що захищає їх від втрати води і висихання. В онтогенезі шкірка диференціюється із зовнішнього шару меристеми конуса наростання. Епідерма, як правило, одношарова. Зрідка у рослин посушливих місць (олеандр, фікус) зустрічається епідерма дво- і тришарова. Епідерма складна тканина. Складається з таких компонентів: 1. основна тканина – клітини якої утворюють велику частину її поверхні; 2. продихи і зв’язані з ними побічні клітини – продиховий апарат; 3. різні вирости (волоски). Клітини основної тканини епідерми паренхімні, щільно зімкнені, іноді мають звивисті стінки, що сприяє міцнішому їх з’єднанню. Епідерма органів, витягнутих в довжину (стебло, черешки) складається з клітин, подовжених паралельно поздовжньої осі органа. Бічні та внутрішні стінки клітин тонкі целюлозні, а зовнішня потовщена і вкрита кутикулою. Часто зверху кутикули розташовується восковий наліт, який, як і шар кутину, зменшує випаровування води з поверхні органа, а у рослин вологих місць перешкоджає змочуванню водою, накопичення якої на листках може сприяти розвитку грибів та бактерій. Основні клітини епідерми живі. З пластид хлоропласти зустрічаються рідко, в основному у водних рослин. Проте, сьогодні деякі вчені визнають, що клітини основної епідерми, в більшості випадків, містять не лейкопласти, а хлоропласти з слабо розвиненою системою мембран, з низьким вмістом хлорофілу і низькою фотосинтетизуючою властивістю. Газообмін між внутрішніми тканинами і зовнішнім середовищем, а також випаровування води здійснюється через продихів. Продих – це щілина, оточена двома замикаючими клітинами. Дані клітини мають бобоподібну форму та нерівномірно потовщені целюлозні оболонки. Стінки, що прилягають до продихової щілини потовщені, а інші – тонкі. На поперечному зрізі щілина між замикаючими клітинами розширена воронкоподібно назовні і в середину органу. Ці розширення називаються переднім і заднім двориком. Оболонки замикаючих клітин вкриті кутикулою. Замикаючі клітини живі, мають протоплазму з хлоропластами та великою кількістю мітохондрій, а також велику вакуолю.
У деяких рослин поруч замикаючими розташовані так звані побічні клітини, які відрізняються за формою від основних клітин епідерми. Вважають, що вони сприяють транспорту води у замикаючі клітини. Замикаючі і побічні клітини утворюють продиховий апарат. Рослини здатні регулювати кількість води, що випаровується збільшенням або зменшенням розмірів продихової щілини шляхом її відкривання і закривання. Продихові рухи відбуваються осмотичним шляхом. Замикаючі клітини активо (з витратою енергії) надходять із зовнішніх клітин іони калію, що веде до зростання осмотичного тиску в їх вакуолях. Внаслідок цього замикаючі клітини поглинають воду, збільшується їх об’єм та тургор, тонкі стінки розтягуються і тягнуть за собою потовщені – продих відкривається. Відтік іонів калію з вакуолей замикаючих клітин відбувається пасивно (без витрат енергії). При цьому спадає осмотичний тиск у вакуолях, клітини втрачають воду. Напруга тонких стінок зменшується, потовщені стінки зближуються – продих закривається. Рух продихів регулюється також світлом, забезпеченістю рослини водою, концентрацією вуглекислого газу в міжклітинниках. Замикаючі клітини утворюються з однієї ініціативної клітини продиху, яка ділиться на дві дочірні. Між ними в результаті часткової мацерації (розчинення) виникає щілина. Під продихом розташовується повітр’яна порожнина, яка сполучається з внутрішніми міжклітинниками. Продихи розміщені на всіх надземних органах рослин, але особливо багато їх в епідермі листкових пластинок (кілька сотень на 1 мм2). У дводольних продихи безладно розкидані в епідермі. У однодольних вони розміщені правильними рядами. У рослин, які ростуть в посушливих умовах продихи розташовані у заглибинах, а саме, замикаючі клітини знаходяться нижче рівня основних клітин епідерми.

Рис. 1. Будова продиху. У багатьох рослин клітини епідерми утворюють вирости, волоски або трихоми у вигляді горбків, сосочків, щетинок, зірчастих лусочок, кущиків (у яблуні, хмелю, фіалки, підмареника, корв’яка). Волоски створюють опушеність органів рослин. Опушеність листка може бути різною. Як правило, більше волосків розвиваються на нижньому боці листка. На стеблі волоски іноді розміщені рядами. Волоски можуть бути одноклітинними – не відокремлюються перегородкою від клітин епідерми, з яких вони утворились, і багатоклітинними. Вміст їх клітин часто відмирає і вони заповнюються повітр’ям. Такі волоски мають білий або сіруватий колір, які відбивають сонячні промені і таким чином захищають рослину від перегріву та зайвого випаровування. Довгі, тверді, густо розташовані волоски перешкоджають поїданню рослин комахами-шкідниками, а іноді й травоїдними ссавцями. Іноді з верхньої клітини багатоклітинних волосків виділяються специфічні речовини. В цьому випадку трихоми називають залозистими. Також бувають жалкі волоски.
Від трихом, що виникають з клітин епідерми відрізняються емергенци (шипи), які зустрічаються на поверхні пагонів, черешка, на жилках листків та деяких інших органах рослини. У деяких рослин (малина, ожина) вони складаються з сукупності клітин, оформлених у тверді, загострені структури. Емергенци утворюються внаслідок поділу та зростання клітин не лише епідерми, але і паренхімних клітин, які лежать під шкіркою.
З ростом і розвитком рослин епідерма змінюється вторинною покривною тканиною – перидермою. Наприкінці літа клітини шкірки стебла або живі клітини, які лежать під шкіркою (субепідермальний шар) набувають здатності до ділення і перетворюються у вторинну твірну тканину – фелоген (корковий камбій). Клітини фелогену діляться тангентально (паралельно поверхні) і відкладають до периферії численні шари корку або фелеми, а в середину 1-2 шари живої тканини фелодерми. Всі ці три шари (фелема, фелоген, фелодерма) називаються перидермою. Власне покривну функцію в цьому комплексі виконує корок (фелема), а корковий камбій, який складається з одного шару клітин, постійно його продукує. Корок складається з правильних радіальних шарів щільно зімкнутих клітин, в оболонках яких відсутні пори. Стінки клітин просочені суберином, корковіють, вміст їх відмирає і заповнюються повітр’ям, що надає корку властивості слабкої теплопровідності. Корковий покрив захищає рослину від різкої зміни температури, проникнення мікроорганізмів, висихання. Клітини корку можуть заповнюватись смолами і набувати коричневого або жовтого забарвлення. У берези в порожнині клітин корку відкладається біла речовина бетулін, за рахунок чого стовбур берез має білий колір. Товстий шар корку дуба відрізняється еластичністю та як термо- і звукоізоляційний матеріал.
Рис. 2. Схема будови перидерми. 1 – залишки епідерми, 2 – фелема, 3 – фелоген, 4 – фелодерма, 5 – пухка паренхіма первинної кори.
Газообмін і транспірація корку здійснюється через сочевичку, що приходить на зміну продиха в процесі утворення перидерми. Під групою продихів починають ділитись клітини, утворюючи пухку тканину з круглястих, тонкостінних, безхлорофільних клітин, яка піднімає шкірку і розриває її. Виникає заглибина у вигляді кратера, оточеного валиком – сочевичка. Клітини, які заповнюють сочевичку внаслідок мацерації втрачають зв’язок одна з одною, округлюються, часто корковіють, утворюючи заповнюючу (або виповнюючу) тканину. Через неї відбувається провітрювання внутрішніх тканин рослини. Під сочевичкою закладається фелоген, що змикається з фелогеном перидерми, і продукує нові клітини виповнюючої тканини. Напередодні зими фелоген сочевички формує замикаючий шар корку, який захищає рослину від висихання. Проте, дерева навіть взимку не бувають закриті герметично, а саме, у замикаючому шарі є невеликі, вузькі міжклітинники, через яких відбувається мінімальний газообмін. Навесні замикаючий шар розривається під напором виповнюючої тканини. Сочевички виступають над поверхнею корку у вигляді різноманітних горбиків, які можуть бути круглястими, ромбоїдними, довгастими. З часом форма і розміри сочевичок може змінюватись. У деяких дерев (осика, бук, ліщина) перидерма зберігається до кінця життя. З середини фелоген відкладає все нові і нові шари, а ззовні клітини корку розриваються і злущуються. При цьому поверхня стовбура залишається гладкою.
Рис. 3. Будова сочевички. 1 – перидерма, 2 – первинна кора, 3 – фелоген сочевички, 4 – коленхіма, 5 – виповнюючі клітини, 6 – новоутворені виповнюючі клітини
Перидерми можуть закладатись концентричними колами, при цьому зовнішні шари кірки тріскаються і злущуються у вигляді поздовжніх смуг. Така кірка називається кільчастою (виноград, кипарис, евкаліпт). Однак, частіше нові перидерми закладаються увігнутими дугами, а не суцільним кільцем. При цьому зовнішні шари кірки злущуються у вигляді лусок (дуб, в’яз, платан). У яблуні кірка виникає в 6-8 річному віці, у сосни у 8-10, у дуба в 25-30, у граба 50 річному віці. Для рослин кірка є надійним захистом від перегріву та опіків у лісових пожежах.
МЕХАНЧНІ ТКАНИНИ
Після виходу рослин на сушу в процесі еволюції в них виникли спеціальні механічні тканини, які підтримують тіло рослин, надають йому форми та положення у просторі. Завдяки механічним тканинам стало можливим збільшення розмірів тіла рослин. Основною особливістю механічних тканин, пов’язаною з їх функцією, є наявність потовщених клітинних оболонок, які часто здерев’яніють. Механічні тканини називаються ще опорними або арматурними.
Розрізняють два види механічних тканин: коленхіму та склеренхіму.
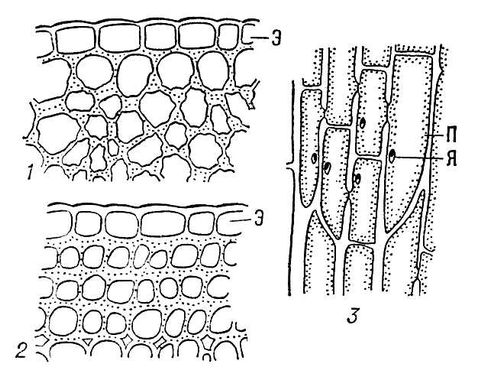
Коленхіма складається з більш-менш видовжених клітин. Клітини живі, містять всі компоненти протопласта, хлоропласти, центральну вакуолю. Клітинні стінки целюлозні, мають нерівномірні потовщення. За типом потовщень виділяють 3 групи коленхіми: 1. кутову; 2. пластинчасту; 3. пухку.
Стінки кутової коленхіми потовщені в кутах, де сходяться кілька клітин (стебла гречки, гарбуза, щавлю). В клітинах пластинчастої коленхіми потовщені тангентальні стінки (соняшник), а у пухкої – стінки біля міжклітинників. Нерівномірне потовщення оболонок не перешкоджає ріст клітин за рахунок непотовщення ділянок росту органів. Коленхіма, звичайно, зустрічається у молодих органах дводольних рослин. У однодольних коленхіма відсутня.
Рис. 4. Коленхіма. 1 – кутова, 2 – пухка, 3 – пластинчаста, Э – епідерма, П –протопласт, Я – ядро.
Склеренхіма – основна механічна тканина рослин. Характерна як для одно-, так і для дводольних. Клітини мертві з товстими здерев’янілими оболонками та з товстими порами. Склеренхіму поділяють на 2 групи: 1. волокна; 2. склереїди.
Склеренхімні волокна являють собою довгі прозенхімні клітини із загостреними кінцями. Стінки їх потовщені, здерев’янілі, а саме, діаметр внутрішньої порожнини може бути меншим за діаметр товстої оболонки. Пори прості, щілиноподібні, розміщені під кутом до поздовжньої осі клітини. Живий вміст волокон рано відмирає. Склеренхімні волокна – міцна тканина, яка за міцністю приближається до сталі. По рослині волокна проходять пучками, в яких окремі клітини закінчуються на різному рівні, перекриваючи одна одну, що підвищує міцність тканини. Склеренхімні волокна, що проходять в лубі називаються луб’яними, а ті, що проходять в деревині – деревинними, або лібриформом. Луб’яні волокна значно довші за лібриформом та більш потовщені. У волокон лубу стінка клітин целюлозна або трохи здерев’яніла. У волокон деревини стінка здерев’яніла, проходить лігніфікацію. Волокна можуть розташуватись не тільки поблизу провідних тканин, але і в первинній корі стебла під шкіркою у вигляді суцільного кільця або окремих тяжів, а також у жилках листків. Луб’яні волокна ряду рослин використовуються як сировина для текстильної промисловості (льон, коноплі). При цьому цінується довжина волокна і мале їх здерев’яніння. Слабкоздерев’янілі волокна використовують для виготовлення високоякісних тканин. Волокна коноплі більш здерев’янілі і вони ідуть на виготовлення грубих тканин.
Рис. 5. Склеренхімна. 1 – окремі склеренхімні клітини, 2 – пучок склеренхімних клітин, 3 – склеренхіма на пеперечному зрізі.
Склереїди або кам’янисті клітини – частіше паренхімні, рідше витягнуті та розгалужені. Стінки їх потовщені, здерев’янілі, мають прості округлі пори, канали яких можуть розгалужуватись. Клітини мертві. За формою клітин склереїди поділяються на: 1. брахісклереїди – вони, як правило, паренхімні; 2. астросклереїди – це потовщені, розгалужені клітини.
Рис. 6. Склереїди - кам’янисті клітини.
Брахісклереїди розташовуються по одинці у тканинах плода, стебла (дрік), кореневищах (півонія), коренях (хрін). Вони можуть утворювати щільну кам’янисту тканину (шкаралупа горіха, кісточки вишні, абрикоса).
Рис. 7. Різні типи склереїд. а – брахісклереїди, б – астросклереїди Астросклереїди – великі розгалужені опорні клітини. Зустрічаються у вигляді ідіобластів у м’якуші листка (чай, малина), а також у тканинах водних рослин, що мають великі міжклітинники (латаття, глечики).
ПРОВІДНІ ТКАНИНИ
Після виходу рослин на сушу в процесі пристосування до нових умов життя виникали два середовища живлення: ґрунтове (мінеральне) і повітр’яне (фотосинтетиз). Виникла необхідність пересувати воду та інші речовини на великі відстані у двох протилежних напрямках. З коренів угору вода і розчинені в ній мінеральні речовини – висхідна течія – рухаються по ксилемі. З листків вниз органічні речовини – низхідна течія – рухаються по флоемі. Функцію проведення речовин по рослині виконують спеціальні провідні тканини. Водний розчин мінеральних і деяких органічних речовин із коренів у листки пересувається по ксилемі, а органічні продукти фотосинтезу з листків по флоемі. Розчин речовин, який транспортується по провідних тканинах називається пасокою. Провідні тканини мають ряд спільних ознак: 1. Обидві тканини формуються з тяжів прокамбію і камбію. При цьому з прокамбію утворюється первинна ксилема і флоема, а з камбію вторинні ці елементи. 2. При диференціюванні провідних тканин з прокамбію на початку онтогенезі спочатку виникає протоксилема і протофлоема, а пізніше метексилема і метафлоема. 3. Вторинна ксилема і флоема складається тільки з метаелементів. 4. Елементи обох провідних тканин мають видовжену форму, що зв’язано з їх провідною функцією, і численні пори – перфорації – які полегшують пересування речовин. 5. Ксилема і флоема складні тканини, оскільки до їх складу крім провідних елементів входять і механічні волокна та паренхімні клітини. Однак провідні елементи в цих тканинах є головними, тому що саме в них відбувається пересування речовин по рослині. 6. Провідні тканини знаходяться поруч, тісно взаємопов’язані і тягнуться по всій рослині від кінчика кореня до верхніх молодих листків, утворюючи провідну систему.
Ксилема являє собою мертві елементи зі здерев’янілими стінками – трахеї та трахеїди. Їх називають ще трахеальними елементами. Трахеїди – це прозенхімні клітини, що сягають від 1-4 до 10 мм довжину, у ширині від 10 до 100 мм, з косо зрізаними, загостреними або заокругленими кінцями. Косі стінки мають численні пори, через яких іде пересування речовин. Пори є і на бічних стінках, через яких відбувається фільтрація води в радіальному напрямі від однієї трахеїди до іншої, а також між трахеїдами та сусідніми паренхімними клітинами. В процесі диференціювання прокамбіальні та камбіальні клітини ростуть з своїми кінцями, де соререджена основна маса органел протопласта, в тому числі пухирці апарату Гольджі, які беруть участь в утворенні клітинної оболонки. Коли ріст клітин та формування їх оболонок закінчується, в протоплазмі з’являються багато лізосом, гідролітичні ферменти яких здійснюють автоліз – розчинення живого вмісту. В порожнину відмерлих клітин надходить вода і з її течією виносяться залишки зруйнованого протопласта. Трахеїди починають функціювати як провідні елементи. За характером потовщення клітинної оболонки трахеїди поліляються на три типи: 1. кільчасті; 2. спіральні; 3. пористі. У кільчастих трахеїд вторинна оболонка накладається на первинну не по всій поверхні клітини, а у вигляді кілець різної ширини. Кільчасті потовщення при нестачі води перешкоджають сплющенню поздожних стінок трахеїд, хоча і не заважає їх деякого прогинанню в середину. У спіральних трахеїд вторинна оболонка накладається на первинну у вигляді спиралі, що є надійнішою опорою для клітинної стінки ніж окремі кільця. Іноді зустрічаються трахеїди, які мають вторинне потовщення у вигляді спіраль і кілець. Кільчасті та спіральні трахеїди притаманні протоксилемі. У них велика частина площі клітинної стінки складається тільки з первинної оболонки, яка здатна розтягуватись, що є важливим, оскільки протоксилема диференціюється в органах рослини які активно ростуть. При розтягненні трахеїд відстань між кільцями збільшується, а кут нахилу витків зменшується. З рештою елементів протоксилема руйнується. Залишки зруйнованої протоксилеми сплющуються під тиском навколишніх клітин, а у стеблах деяких однодольних рослин на місцях зруйнованих трахеїд залишаються порожнини – протоксилемні лакуни. Протоксилема має простішу будову у порівнянні з метаксилемою. Крім трахеальних елементів містить паренхімні клітини, а механічні елементи відсутні. Пористі трахеїди є компонентами первинної і вторинної метаксилеми. Своє формування вони закінчують в органах, які припинили ріст. Вони не здатні до розтягування і зберігаються протягом усього життя рослини, хоча згодом втрачають свою провідну функцію. Типи поровості трахеїд метаксилеми різні. Розрізняють поровість драбинчасту, супротивну і безладну. Трахеїди – найдавніші водопровідні елементи рослин (особливо спіральні та кільчасті). У вищих спорових та голонасінних рослин течія води відбувається у трахеїдах. У покритонасінних також зустрічаються трахеїди, але в основному в протоксилемі. Присутність трахеїд в метаксилемі свідчить про давність і певну примітивність даного виду.
Рис. 8. Елементи трахеїдальної провідної тканини. а – волокно з коротких запасаючих клітин, б – запасаючі клітини, в – трахеїда, г – клітина механічної тканини, д – тонкостінна трахеїда, е – товстостінна трахеїда.
Трахеї або судини – це трубки, які складаються з вертикального ряду мертвих клітин-члеників, що мають перфорації на поперечних стінках. Вони є більш удосконалими елементами, оскільки вода безпосередньо перетікає з членика в членик через великі отвори. Членики судин в процесі еволюції рослин утворились з трахеїд, які поступово ставали коротшими і ширшими за рахунок зменшення клітин прокамбію і камбію. В онтогенезі з вертикального ряду камбіальних клітин диференціювались членики. Розвиток члеників схожий на розвиток трахеїд. Відмінність полягає в тому, що потовщені лише поздовжні стінки, в яких утворились облямовані пори. Поперечні ж ослизнюються і набрякають за участю пухирців апарату Гольджі, які накопичуються на кінцях членика, що розриваються. При рості члеників в ширину ослизні стінки розчиняються і утворюють перфорації. Припускають, що в руйнуванні цих оболонок беруть участь і гідролітичні ферменти лізосом і вакуолей, що розщеплюють речовини клітинної стінки. У більшості покритонасінних протоксилема складається з трахеїд, а метаксилема з трахей. Однак, у деяких рослин (бобові, складноцвіті) протоксилема також формується з трахей кільчастих або спіральних, що вважається ознакою більш високої організації. Трахеї довші за трахеїди, можуть сягати декілька метрів. Вони ширші за трахеїди. У деревних покритонасінних трахеї складаються з багатьох тисяч коротких і широких члеників. На думку вчених, трахеї в процесі еволюції дали початок не тільки судинам, але і механічним елементам ксилеми, а саме – лібриформу. У цьому випадку зміни були спрямовані в бік більш значного потовщення оболонки та зменшення кількості пор, що втрачали облямів і ставали щілиноподібними. Проміжними елементами між трахеїдами та лібриформом є трахеальні волокна, які мають менше пор з слабко вираженою облямівкою. Трахеальні елементи ксилеми виконують провідну функцію. По них від коренів угору в рослині відбувається течія води і поглинутих з ґрунту мінеральних речовин. Крім того, у пасоці ксилеми міститься певна кількість органічних сполук: продукти синтетичної діяльності коренів (амінокислоти, фітогормони), продукти гідролізу запасних речовин (амінокислоти, цукри). Трахеальні елементи виконують також опорну функцію завдяки твердим здерев’янілим оболонкам. Паренхімні клітини здійснюють запасну функцію і проводять воду та розчинені в ній речовини в радіальному напрямі. У однорічних рослин ксилема функціонує один рік та відмирає разом з рослиною. У багаторічних з віком у ксилемі відбуваються різні зміни. Трахеальні елементи можуть виконувати свою функцію протягом багатьох років (від 1-2 до 40-50), а потім втрачають її і заповнюються повітр’ям або деякими іншими речовинами (вуглекислий або сірчанокислий кальцій, дубильні речовини, ефірні олії, смоли). Тили – це вирости живих клітин, які проникають в судини, рідше в трахеїди, через їх пори. У вирости переходять ядро, або одне з ядер після його поділу, і центральна вакуоля. Поділ ядра може відбуватись і в самій тилі. Тоді між дочерними ядрами формуються перетинки, і таким чином виникає багатоклітинне утворення – псевдотканина. У тилах відкладаються поживні речовини (крохмаль, олії) та продукти життєдіяльності рослини (дубильні речовини, смоли), які захищають деревину від пошкоджень бактеріями, грибами, комахами. Крім того, тили збільшують міцність ксилеми. Заростання судин тилами відбувається у старих стеблах деревних рослин, рідше в молодих, ще рідше в трав’янистих (гарбузові). У деяких рослин тили не утворюються і закупорення судин відбувається відкладанням мінеральних та органічних речовин (клен, береза, вишня). Вікові зміни в ксилемі спричиняють відмирання живих клітин. При цьому в паренхімних клітинах попередньо потовщується та здерев’яніє клітинна оболонка (склерифікація), і живі клітини перетворюються на механічну тканину. Склерифікація посилює механічну міцність органів рослин.
Рис. 9. Послідовні стадії формування трахеї. 1 – поздовжній ряд меристематичних клітин, 2 – клітини витягувались в довжину і вакуолізувались, 3 – клітини ще більше подовжувались, стінки їх потовщувались, 4 – клітини збільшувались у поперечному напрямі, поперечні стінки ослизнювались і набрякали, 5 – в перегородках утворились перфорації, у поздожніх стінках сформувались облямовані пори, живий вміст клітин утрачено, трахея готова до функціонування.
Флоема. Головними її структурними елементами є ситовидні трубки і клітини-супутники. Ситовидні трубки – це вертикальний ряд живих клітин-члеників, поперечні стінки яких мають перфорації (ситичка). Оболонки двох сусідніх клітин пронизані отворами і називаються ситоподібними пластинками. Отвори на них звичайно розташовані групами, утворюючи ситоподібні поля. У перфораціях проходять тяжі цитоплазми, які з’єднують проторласти сусідніх клітин. Членики ситоподібних трубок мають тонкі целюлозні оболонки і живий вміст, що відрізняється від протопластів звичайних живих клітин. Вони мають плазмолему, що оточує цитоплазму та вистилає перфорації, гладкий ендоплазматичний ретикулюм, тонопласт у них відсутній, клітинний сік змішується з цитоплазмою. Відсутні ядра, пластиди, мітохондрії, апарат Гольджі, рибосоми. Всі ці особливості разом з широкими цитоплазматичними тяжами в перфораціях є пристосуванням до транспорту досить концентрованого (до 90 %) розчину асимілятів. До їх складу входять цукри (80-90 %), амінокислоти, органічні кислоти, фітогормони і вітаміни. На ситоподібних пластинках та на стінках перфорацій накопичується особливий полісахарид – калоза, що спричиняє поступове зменшення отворів. Поруч кожним члеником розташовується одна або декілька клітини-супутники. Це звичайні живі клітини з целюлозною оболонкою і протопластом з усіма органелами. Клітини-супутники забезпечують надходження в ситоподібні трубки продуктів фотосинтезу з клітин м’якуша листка та вихід їх у місцях споживання рослиною (в точках росту, запасаючих органах). В одних клітин-супутниць є надходження асимілятів з фотосинтезуючих клітин через плазмодесми, зібрані у плазмодесм’яні поля, а в інших клітинні оболонки утворюють протуберанці, що збільшують поверхню плазмолеми, через яку транспортуються органічні речовини, при чому основна маса і плазмодесм і протуберанців в клітинах-супутниках розташована у стінках, що прилягають до клітин мезофілу. Зв’язок клітин-супутниць з ситоподібними трубками дещо інший. З боку супутниць в оболонках є плазмодесм’яні поля, що зв’язуються з ситоподібними полями трубок. В тонких жилках листка, де відбувається інтенсивний перехід продуктів фотосинтезу у флоему (завантаження флоеми), у кожному членика ситоподібної трубки є кілька великих клітин-супутниць, при чому їх діаметр часто більший за діаметр ситоподібних елементів. В осьових органах, де в основному здійснюється вихід асимілятів (розвантаження флоеми), клітина-супутниця ситоподібного членика звичайно одна, а якщо їх декілька, то вони утворюють однорядний вертикальний тяж біля його поздовжньої стінки. Елементи флоеми, як і трахеальні елементи, формуються з прокамбію і камбію. Меристематичні клітини поділяються поздовжними перегородками на дві нерівні клітини, а саме одна клітина вужча за іншою. Клітини ростуть, при чому збільшує клітина-членик ситоподібної трубки та зменшує клітина-супутниця, що може далі ділитися поперечною перегородкою і може утворювати тяж уздовж членика ситоподібної трубки. На початку свого розвитку клітина-членик має в протопласті всі органели, вакуолю та численні плазмодесми в клітинних стінках. В процесі диференціювання в цитоплазмі з’являються кільця з Ф-білка. Їх ще називають слизовими кільцями. Поступово слизові кільця розпливаються і набувають нитчасту структуру. До кінця формування члеників нитки Ф-білка розташовуються уздовж плазмолеми та цитоплазматичних тяжах перфорацій. Існує думка, що фібрили Ф-білка здатні до скорочення і допомагають пересуванню асимілятів у ситоподібних трубках. Одночасно з формуванням ниток Ф-білка відбувається руйнування протопласта, вакуолярний сік змішується з цитоплазмою; після формування цитоплазматичних колів руйнується й апарат Гольджі, пластиди, мітохондрії, рибосоми, останнє руйнується ядро. Незважаючи на значні структурні зміни протопласта, клітини-членики ситоподібних трубок залишаються живими, мають напівпроникну плазмолему, гладкий ЕПР і зберігають здатність до плазмолізу. В онтогенезі першою утворюється протофлоема, що складається з вузьких ситоподібних трубок, а клітини-супутники у них часто відсутні. Елементи протофлоеми функціонують недовго і сплющуються під натиском тканин, що ростуть. Пізніше формуються елементи метафлоеми, кожен членик ситоподібної трубки яких супроводжений клітиною-супутницею. Клітинні членики більш широкопросвітні, ширшими є і перфорації ситоподібних пластинок. Ситоподібні трубки значно коротші за судини ксилеми. Довжина їх коливається від 100 до 300 мкм, а ширина від 20 до 30 мкм. Ситоподібні трубки є високоспеціалізованими провідними елементами, що сформувались у процесі еволюції та функціонують у квіткових рослинах. Їх попередниками, напевно, були ситоподібні клітини, що нині присутні у вищих спорових. Це вузькі, довгі клітини із загостреними кінцями, де у первинних оболонках є численні плазмодесми, зібрані у групи – плазмодесм’яні поля. В процесі розвитку плазмодесм’яні канальці часто перетворюються на скрізьні отвори.
Рис. 10. Ситоподібні трубки на поздовжньому (1) та на поперечному (2) зрізі. а – клітини-супутники, б – поперечна перетинка, в – пори.
Еволюція флоеми відбувалася в напряму скорочення ситоподібних клітин, збільшення їх діаметра та діаметра перфорацій. Поперечні стінки набували більш близьке до горизонтального положення і на них зосереджувалась основна маса ситоподібних полів. У голонасінних функціонують ситоподібні трубки, але відсутні клітини-супутники. Їх роль виконують паренхімні клітини – клітини Страсбургера. Зовні вони не відрізняються від клітин-супутниць, відмінність полягає лише в їх походженні. Клітини Страсбургера і членики стоподібних трубок утворюються з різних меристематичних клітин. Флоема – складна тканина. Крім ситоподібних трубок і паренхімних клітин містить і луб’яні волокна. Луб’яні волокна використовують у текстильній промисловості, при чому особливо цінується їх довжина і відсутність здерев’яніння. Флоема – тканина недовговічна. Ситоподібні трубки функціонують один сезон, рідше 1-2 роки. У однодольних ситоподібні трубки функціонують довше ніж у дводольних. Нетривалість життя провідних елементів флоеми пояснюється відсутністю ядра в клітинах-члениках та накопиченням калози. Згодом шар калози на ситоподібних пластинках збільшується, а перфорації перетворюються у вузькі канальці. До кінця вегетативного періоду канальці зовсім закупорюються, а на ситоподібній пластинці утворюється наплив – калюс, мозолисте тіло. Ситоподібні трубки, що припинили функціонувати відмирають і під напором сусідніх тканин сплющуються. Слідом з ситоподібними трубками відмирають і клітини-супутники.
ПРОВІДНІ ПУЧКИ
|
||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-18; просмотров: 1429; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.197.123 (0.112 с.) |