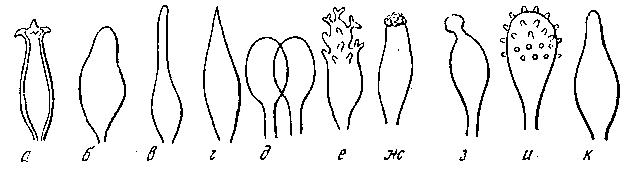Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Человек и окружающая среда –Содержание книги Поиск на нашем сайте Лидия Григорьевна Бурова Загадочный мир грибов
Человек и окружающая среда –
Л. Г. Бурова Загадочный мир грибов
Памяти В. Н. Сукачева и Н. В. Дылиса
АКАДЕМИЯ НАУК СССР Ответственный редактор М. В. Горленко
От автора
В этой книге речь пойдет о грибах, о тайнах их жизни. Так называемые высшие грибы, выделяемые в последнее время вместе с «низшими» в отдельное царство, оказались сейчас в невыгодном положении. С одной стороны, многие знают, где, когда и в каких лесах их можно много собрать, какие грибы нельзя собирать (они ядовиты и ими можно отравиться), с другой – даже биологи мало осведомлены о них. Исключение составляет небольшое количество специалистов, называемых микологами. Их у нас в СССР немногим более 100 человек. Все грибы: и те, о которых пойдет речь в этой книге, и те, которые можно увидеть только под микроскопом, – совершенно удивительные представители жизни на Земле. До сих пор о происхождении грибов и их эволюции мы знаем очень мало. Их можно с полным правом назвать космическими пришельцами. Знать необходимые для каждого человека сведения о грибах нужно потому, что в самом ближайшем будущем стремительно возрастет вмешательство грибов в нашу жизнь, так же стремительно, как возрастает техногенное воздействие на окружающую нас природу. Мне, как автору этой книги, очень хотелось, чтобы читатели имели ясное представление о том, что же такое любимые нами грибы? Как и где они живут, где их можно собирать, а где даже такие благородные их представители, как белые, грузди, рыжики, могут вызвать тяжелейшие отравления; какую пользу и какой вред могут нанести людям объекты «тихой охоты»? Знаете ли вы, например, что свинушка, о которой очень много писали, оказывается токсичной только в местах загрязнения промышленными отходами, вдоль шоссейных и железных дорог, а также в городах. Это касается не только свинушки, но и шампиньонов и других грибов, собираемых в парках, садах городов, всех овощей и фруктов, произрастающих в подобных местах. Грибы – индикаторы различной степени антропогенной нагрузки на леса. Наличие, например, большого количества и видового разнообразия сыроежек указывает на то, что вытаптывание в березняках Подмосковья достигло предельно допустимых норм. Грибы (18 видов) занесены в настоящее время в Красную книгу СССР. Они нуждаются в охране: съедобные и ядовитые, а в особенности те, которые имеют причудливые и декоративные формы и окраску. В нашей стране на душу населения приходится около 30 кг/год грибов, однако потребляется менее 1 кг/год. Остальные грибы первой категории остаются несобранными в лесах. Для того чтобы мы находились на среднем уровне потребления грибов в Европе, необходимо рациональное размещение грибозаготовочных пунктов. Это мероприятие не требует больших материальных затрат. Первая заповедь грамотного «грибника» – не собирать не знакомых Вам грибов, вторая – не собирать грибы, ягоды, плоды в местах, подверженных загрязнению промышленными отходами; третья – внимательно читать книги о грибах, а не просматривать только «картинки». В написании этой книги неоценимую помощь оказали родные и близкие для меня люди. Их очень много! Нет надобности перечислять их поименно, но особую благодарность я выражаю моему сыну А. Г. Бурову, докторам биологических наук Н. М. Черновой, А. И. Уткину, кандидатам биологических наук Э. Л. Нездойминого, П. В. Гордиенко, М. М. Гьошевой‑Богоевой, старшим лаборантам А. Н. Кузнецову, А. Ю. Семашко и др.
В царстве грибов
Среди огромного разнообразия жизни на Земле мы видим во все времена года на деревьях копытообразные наросты, а на валежных стволах с ранней весны и до поздней осени «живут» мхи, лишайники и чрезвычайно разнообразный мир грибов. К настоящему времени насчитывается около 100 тыс. видов высших грибов, но и это не предел! Вполне вероятно, что их гораздо больше, так как ежегодно описывают новые виды и формы. Не следует забывать, что большая часть суши и моря совсем не исследованы «флористически» (т. е. здесь не выявлен видовой состав грибов). Во многих районах Вьетнама, Кореи, Лаоса и других регионах после ведения в них войн возникло огромное количество мутантов, а поскольку войны на земле до сих пор не прекращаются, можно себе представить, что при применении современных видов оружия количество новых видов грибов будет неуклонно возрастать. Поэтому необходимо в кратчайшие сроки всем людям Мира сесть за круглый стол и проанализировать, до чего же человечество довело нашу землю, и, естественно, принять срочные меры по спасению оставшегося пока органического мира на нашей земле. Коснемся кратко истории происхождения и изучения грибов. Грибы расселены по всему земному шару. Споры их в бесчисленном количестве заполняют воздух, воду и землю. Богаче всех грибами жаркие тропические страны (влажные тропические леса). Однако лучше изучены в микологическом отношении территории стран умеренного климата (Европа, Северная Америка). В странах Западной Европы обнаружено более 5 тыс. видов грибов. Большинство грибов невидимы простым глазом, и только сильное увеличение микроскопа позволяет нам увидеть их, проникнуть в тайны их жизни. Оказалось, что у всех грибов одинаковый принцип строения. Плодовые тела грибов «прорастают» не из семени, как это считалось ранее, а из мельчайших спор, поэтому они относятся к споровым организмам, как, впрочем, мхи и лишайники. Если у цветковых растений в семени всегда можно различить зачатки стебля, листа и корня, то у грибной споры ничего этого нельзя обнаружить, так как она представляет собой обычно обыкновенный «пузырек, заполненный жидкостью». Такая спора, попадая в подходящие для нее условия обитания, питаясь подходящим для нее субстратом, начинает постепенно развиваться, т. е. «прорастать». Споры по форме, структуре, цвету могут быть самыми различными (рис. 1). Многообразие форм спор может дать отдаленное представление о разнообразии самих грибов. Стоит лишь войти в лес и внимательно взглянуть вокруг себя, как вы увидите, какие могут быть грибы, удивительные, неповторимые, не похожие ни на кого!
Рис. 1. Форма спор а – угловатая; б – веретеновидная; в – шаровидная; г, д – ребристая; е – фасолевидная; ж – лимоновидная; з – каплевидная; и – сетчатая; к – бугристая; л – шиповатая; м – гладкая; н – бородавчатая
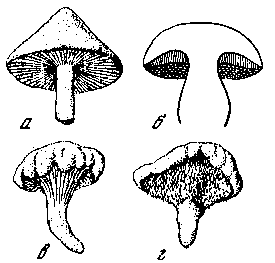
Макромицетами называют группу высших грибов, имеющих крупные плодовые тела различной формы (шляпковидная, шаровидная, в виде звезд, кустиков и т. д.). Они относятся к двум классам грибов – базидиальным (Basidiomycetes) и сумчатым (Ascomycetes). Обычно грибом называют плодовое тело (карпофор) макромицетов, не учитывая того, что основная, рабочая часть его скрыта от нас под слоем опавших листьев и веток, в гумусовом слое, подстилке, почве, древесине. Иногда мицелий – функциональная часть гриба – расположен внутри опавших иголочек хвойных пород, листьев деревьев и кустарников, внутри упавших стволов и ветвей деревьев, одним словом, мицелий различных грибов пронизывает любой субстрат в лесу, на лугах и полях. Грибница, или мицелий, состоит из тончайших нитей (гифов), переплетающихся в субстрате в виде паутины. По своему происхождению грибница вторична, потому что образуется в результате слияния первичного мицелия, возникшего из двух разнополых прорастающих спор. На определенном этапе своей жизни из гиф мицелия образуется (возникает в месте уплотнения «паутины») крохотный узелок – так называемый примордий, из которого при определенных благоприятных сочетаниях тепла и влаги за сравнительно короткий период развивается плодовое тело гриба. Оно представляет собой плотно сплетенные гифы (ложную ткань). Особенности морфологического и анатомического строения грибов являются основой для их определения, т. е. основой систематики грибов (Жизнь растений, 1976). У многих видов макромицетов (агариковых, части афиллофоровых, пецициевых) плодовое тело состоит из шляпки и ножки. Шляпки очень разнообразны по величине, форме, окраске, размеры их варьируют от нескольких миллиметров до десятков сантиметров в диаметре. Форма шляпки в зависимости от фенологии, характера погоды, местообитания и т. д. меняется по мере роста и развития гриба. У молодых плодовых тел она обычно бывает конической, полушаровидной, яйцевидной или цилиндрической, а с возрастом становится колокольчатой, плоской, воронковидной и т. п. Неодинаков у различных видов и край шляпки. Цвет шляпок очень разнообразен и меняется в зависимости от комплекса факторов местообитания. Иногда вследствие различной интенсивности окраски на поверхности шляпки обособляются концентрические зоны, имеющие важное таксономическое значение. У некоторых видов шляпка пропитана водой и почти прозрачна, такие шляпки называют гигрофанными. Консистенция плодовых тел макромицетов также необычайно разнообразна: от мягкомясистой до деревянистой и кожистой. Мякоть шляпок грибов бывает ломкая, упругая, рыхлая или волокнистая, толстая или тонкая. У мякоти имеется определенный вкус (пресный, сладкий, горький, кислый, острый, жгуче‑едкий и т. д.) и запах – фруктовый, мучной, редечный, селедочный, кумариновый и т. п. У представителей рода груздей в мякоти содержится млечный сок – сладковатый или жгуче‑едкий. На воздухе его окраска изменяется, становясь лиловой, белой, бесцветной; эти особенности являются таксономическими для грибов указанного рода. Окраска мякоти у них неяркая – от чисто белой до желтой, красноватой, серой или фиолетовой.
Рис. 2. Тип гименофора а – пластинчатый; б – трубчатый; в – складчатый; г – шиповатый
Немаловажным систематическим признаком, как и все предыдущие особенности морфологии и анатомии грибов, является нижняя поверхность шляпки – гименофор, который несет на себе функцию кладовой спорового порошка. По форме он бывает трубчатым, как, например, у белого гриба, масленка, подберезовика, или пластинчатым – у рыжиков, сыроежек, волнушек, шампиньонов, рядовок и т. д. У пластинчатого гименофора пластинки бывают свободными или приросшими, низбегающими по ножке, редкими или частыми, прямыми или разветвленными. Если положить свежесрезанную шляпку гриба на листок бумаги (светлый или, наоборот, темный), то через несколько часов можно получить отпечаток спорового порошка и по нему узнать структуру и морфологию гименофора. Пластинки различаются также окраской, изменяющейся с возрастом плодового тела гриба. Трубчатый гименофор, как и пластинчатый, прикрепляется к ножке различными способами (рис. 2). Гименофор трубчатых грибов различается по цвету, размеру и форме пор – отверстий трубочек. Поры бывают округлые, овальные, угловатые, мелкие или крупные. Трубчатый слой у одних видов легко отделяется от мякоти шляпки, а у других снимается с трудом. Складчатый гименофор сходен с пластинчатым, но складки обычно бывают толще пластинок (у лисичек). Существует еще шиповатый гименофор, он состоит из шипиков на нижней поверхности шляпки, его можно увидеть у таких распространенных видов грибов, как, например, ежовик настоящий, коралловый. Спороносный слой – гимений – у базидиальных грибов состоит из базидий со спорами и стерильных элементов, называемых в микологии цистидами; у сумчатых грибов – из сумок со спорами и стерильных апофиз. Споры различаются по форме, характеру поверхности и многим другим признакам (рис. 1). Цистиды, как и споры, характеризуются определенными морфологическими признаками (рис. 3).
Рис. 3. Форма цистид а – гарпуновидная; б – мешковидная; г – ланцетовидная; д – пузыревидная; е – отросчатая; ж – с кристалликами; з – головчатая; и – с выростами; к – бутыльчатая
Спороносный слой у некоторых видов грибов в начале их развития прикрыт тонким пленчатым покрывалом, общим или частным, иногда оно выглядит, как паутинка на концах шляпки и ножке гриба (род паутинники). У зрелых карпофоров покрывало разрывается и сохраняется в виде лоскутков на шляпке (мухоморы), либо на ножке в виде кольца (шампиньоны). У некоторых видов имеется как общее, так и частное покрывало, у других – только одно. Грибница макромицетов, накопив достаточное количество питательных элементов и энергии, образует плодовое тело гриба. Плодоношение, или плодообразование, грибов зависит от целого ряда факторов, но в микологии принято считать, что ведущими факторами при этом процессе выступает гидротермический режим местообитания. Подавляющее большинство макромицетов плодоносит при влажности субстрата не ниже 50%. Избыток влаги, как и ее недостаток, отрицательно влияет на рост и развитие грибов. Сроки плодоношения грибов (фенология) зависят не только от условий внешней среды, но и от биологических свойств и ритма развития самого гриба (биоритма). В зависимости от погодных условий период плодообразования грибов ускоряется или замедляется; если в подходящие для грибов сроки их плодоношения нет необходимых условий, то образование карпофоров задерживается иногда на несколько лет.
Экологические группы грибов
Экологические группы грибов – понятие не таксономическое. В процессе эволюции у грибов, характеризующихся развитым генетическим и биохимическим адаптивным аппаратом, сложились тесные взаимосвязи с автотрофными организмами. Это в основном и определило их пространственное распределение и разделение на экологические группы. Критерии выделения экологических групп до сих пор вызывают многочисленные дискуссии микологов. Пищевые связи макромицетов в лесных сообществах, как правило, определяют их экологические особенности, поэтому в данном случае можно говорить об экологических группах грибов, равнозначных трофическим. Выделяемые отечественными и зарубежными микологами, экологические группы грибов, приуроченные к субстрату или симбиотрофно связанные с растениями без учета местообитания, правильнее было бы назвать трофическими, поскольку экология большинства видов грибов пока что не изучена, и отчасти поэтому до сих пор в микологии нет унификации в названии экологических групп грибов. Трудности в определении экологических групп шляпочных грибов объясняются еще и тем, что большая часть грибов либо мобильна в отношении типа питания (не облигатна), либо, сохраняя его, может образовывать плодовые тела в различных по экологической обстановке условиях, в не типичных для гриба местах. Так, например, обычные в переувлажненных местообитаниях (заболоченные сосняки, березняки, ельники) микоризные грибы – свинушка тонкая, желчный гриб и некоторые другие виды – визуально могут быть отнесены к деревообитающим грибам, так как их плодовые тела встречаются на пнях и кочках, образованных на месте упавших или сломанных деревьев. Однако при этом связи между корнями живых деревьев и мицелием грибов не теряются, поэтому трофически (по типу питания) они, безусловно, относятся к микоризообразующим грибам, а топически (по условиям местообитания) – к деревообитающим (ксилотрофам). В литературе имеются сведения о грибах, способных образовывать плодовые тела наравне с симбиотрофным типом питания сапротрофно (т. е. без связи с корнями деревьев), и таких грибов выявлено немало. По‑видимому, это явление в природе распространено гораздо шире, чем предполагалось ранее. Интересно, что вынуждает гриб менять тип питания: условия местообитания для самого гриба или нарушение взаимосвязей с растениями? По нашим наблюдениям, в неблагоприятных для деревьев условиях, например в сфагновых сосняках и березняках, в лесах, находящихся на границах географического распространения, в искусственных лесных насаждениях, растущих в условиях нелесной зоны (лесозащитные полосы в степях, полупустынях), такие грибы, как свинушка тонкая и лаковица, ведут себя как истинные микоризообразователи. В лесных сообществах, находящихся в оптимальных (благоприятных) для деревьев условиях местообитания, они меняют тип питания на сапротрофный. Только планомерные и длительные наблюдения за грибами в природных условиях могут дать ответ на вопрос, к какой экологической группе они относятся в каждом конкретном случае.
Сапротрофные макромицеты
Сапротрофные грибы объединяют макромицеты, использующиеся в качестве источника пищи. Они осуществляют все процессы жизнедеятельности за счет мертвого органического вещества. Они специализируются на разложении особо стойких лигноцеллюлозных соединений. В процессе эволюции у них сформировался специфический набор ферментов, определивший разделение грибов на ряд экологических групп, основными из которых являются: подстилочные и гумусовые сапротрофы, ксилотрофы, копро‑, карбо‑, брио‑ и микотрофы. Сапротрофы благодаря наличию разветвленной сети гифов обладают уникальной способностью максимального контакта с субстратом, повышающего эффективность метаболических процессов, происходящих в мицелии. Обладая высокой активностью метаболизма, биохимической мобильностью и способностью быстро реагировать на действие неблагоприятных факторов среды переходом к анабиозу, сапротрофы могут осваивать чрезвычайно различные по условиям местообитания экологические ниши. Решающую роль в формировании экологических групп сапротрофных грибов играют биохимические адаптации. Скорость разложения мертвого органического вещества, заключенного в лесном опаде и подстилке, а в связи с этим и интенсивность биологического круговорота веществ в лесных сообществах во многом зависят от биологической активности сапротрофных грибов, выделяющих в субстрат ряд ферментов (целлюлазы, оксидазы, органические кислоты и физиологически активные вещества). Энергия процессов минерализации (высвобождение минеральных веществ из органических) растительных остатков определяется количеством в них легкорастворимых органических веществ и соотношением азота и углерода. Принято считать, что соотношение углерод: азот, равное 8:1–10:1, наиболее благоприятно для развития макромицетов, а при уменьшении этого соотношения скорость деструкции (разложения) органических веществ резко падает и продукты неполного разложения целлюлозы и в особенности лигнина в течение продолжительного времени оказываются законсервированными. Поскольку лесная подстилка разлагается в течение длительного времени, она делится на ряд слоев различной степени деструкции, различающихся между собой биохимически и экологически. Верхний слой, состоящий из полностью сохранивших структуру растительных остатков, характеризуется наибольшим разнообразием биохимического состава, максимальной изменчивостью показателей температуры и влажности, значительно ограничивающими потенциальные возможности сапротрофных грибов разрушать органические вещества. Основу растительных остатков в этом слое составляют гемицеллюлоза, целлюлоза и лигнин, на долю последнего приходится от 20 до 35%, большое количество здесь водорастворимых и зольных соединений, азота и углерода, таннинов, жиров, воскосмол. Разнообразный субстрат активно заселяется бактериями, дрожжами и грибами, использующими для своего питания простые сахара, пектин и белковый азот. Последовательность развития организмов на начальных этапах разложения опада протекает по следующей схеме: вначале субстрат заселяют быстрорастущие бактерии (в основном неспороносные) и некоторые низшие грибы, потребляющие водорастворимые органические соединения, за ними следуют представители сумчатых грибов и несовершенные грибы, потребляющие крахмал, их сменяют базидиальные, в основном сапротрофы‑макромицеты, разлагающие лигнин и целлюлозу. Среди сапротрофных макромицетов наблюдается четкая специализация не только по фракциям опада (листья, хвоя, мелкие опавшие ветки, шишки), но и по видовой принадлежности растений, резко различающихся по химизму и прочности тканей. Это позволяет выделить ряд специфических групп макромицетов на опаде. Одни развиваются на хвое, другие – на опавших мелких ветвях хвойных пород. Для опавших листьев липы и дуба характерны представители рода «негниючники», а для березовых листьев – виды рода Mycena. «Своими» грибами заселены отмершие части трав – различными видами в зависимости от географического положения лесных сообществ и условий местообитания. Все сапротрофы, развивающиеся на опаде, отличаются эфемерностью и образуют плодовые тела через несколько дней после выпадения осадков, при достижении относительной влажности приземного слоя воздуха не ниже 60%. Появление плодовых тел спорадическое и массовое, на 1 м2 лесного опада можно насчитать более сотни мелких (высота ножки 2–4 см, диаметр шляпки – 0,5 см) грибов на тонких хрящеватых ножках. Обычно на такие грибы люди не обращают внимания, ведь они не представляют никакой пищевой ценности. Да, не представляют, но без них невозможен был бы полный процесс разложения мертвого органического вещества в лесах, запасы которого огромны (в среднем в лесах Подмосковья опад составляет 2–3 т/га); без них ежегодно накапливающаяся масса опавших листьев, ветвей, хвои в буквальном смысле слова задушила бы леса! Самый верхний слой подстилки, как и опад, по своему положению в системе слоев лесной подстилки значительно варьирует по химизму вследствие периодичности поступления новых порций опада. Высокое содержание легкодоступных питательных веществ, которое является результатом вымывания и выщелачивания опада дождевыми и талыми водами, вызывает высокую активность поселяющихся здесь в массе темноцветных гифомицетов и некоторых несовершенных грибов. Основными представителями макромицетов этого слоя являются виды рода Mycena – М. sanguinolenta, М. vulgaris, сменяющие друг друга от весны к осени. Численность их, как и сапротрофов на опаде, достигает внушительных размеров (50–70 экз./м2) при благоприятных гидротермических условиях, а размеры плодового тела несколько большие (ножка высотой до 5 см, а диаметр шляпки –до 1 см). В нижележащем слое подстилки, называемом ферментативным, происходит процесс интенсивного разложения целлюлозы и лигнина. Базидиальные макромицеты, участвующие в нем, делятся на две группы по принципу вызываемых ими разрушений органических веществ. К первой относятся виды, обладающие способностью разлагать целлюлозу и лигнин, так как основными ферментными реакциями у них являются гидролиз и окисление. Грибы второй группы разрушают только целлюлозу. Скорость ферментативных процессов определяется воздействием многих факторов, основные из которых – кислотность среды, температура, возраст мицелия грибов, биохимический состав субстрата. Способностью гидролизовать углеводную часть подстилки – целлюлозу, составляющую одну треть ее массы, обладают грибы обеих групп, однако представители первой группы гораздо медленнее расщепляют полимерные углеводы из‑за одновременного разрушения ими лигнина. Внеклеточный фермент – целлюлаза, – ответственный за расщепление углеводов, более активен у грибов, специализирующихся на разложении погребенных в подстилке древесине, шишках, коре. У грибов, разрушающих хвою и листья, целлюлоза носит адаптивный характер, о чем свидетельствует возрастание ее активности на средах с целлюлозой. В разложении лигнина – высокополимерного соединения, химическая природа которого до сих пор до конца не установлена, принимают участие те грибы, которые обладают комплексом ферментов‑трансфераз: виды рода Mycena – М. crocata, М. polygramma, М. rosea, М. galericulata. Ферментативный слой подстилки более однороден экологически, так как гидротермические условия здесь менее контрастны, чем в вышележащем слое, поэтому такого видового разнообразия макромицетов‑сапротрофов, которое наблюдалось в опаде и в верхнем слое, нет. Более или менее однородный биохимически субстрат (полуразложившиеся, потерявшие очертания листья и хвоя) разлагается относительно небольшим набором видов, четко сменяющих друг друга во времени. Значительно разнообразят видовой состав грибов этого слоя макромицеты, приуроченные в своем развитии к погребенным древесине, шишкам, коре, ветвям – субстрату, способному сохранять в течение продолжительного времени постоянную влажность. Под ферментативным слоем подстилки расположен гумусовый, однородный по механическому составу. В нем бесструктурное аморфное органическое вещество тесно связано с минеральной частью почвы. Основная черта гумусового подгоризонта подстилки – минимальная экологическая и биохимическая вариабельность по сравнению с описанными слоями. Гуминоподобные соединения представляют собой продукт гидролиза лигнина, заключенного в органическом веществе. Лигнинные мономеры под влиянием ферментов грибов соединяются с углеводами и продуктами разложения мицелия, образуя гуминоподобные комплексы, близкие по своему строению к истинным гуминовым кислотам. Макромицеты, мицелий которых расположен в гумусовом слое, отличаются постоянством видового состава и относительной независимостью от погодных условий вегетационного сезона. Они обладают более или менее крупными плодовыми телами по сравнению с подстилочными сапротрофами верхнележащих слоев подстилки. Типичными представителями гумусовых макромицетов являются шампиньоны, зонтики, говорушки, волоконницы. Многие виды, относящиеся к этой группе, – обитатели безлесных пространств – шампиньоны, навозники, дождевики и ложнодождевики.
География грибов
Распространение макромицетов на огромной территория Советского Союза исследовано неравнозначно. Очень плохо (на уровне ограниченных районов) изучены грибы севера и центра европейской части СССР, республик Средней Азии, Западной и Восточной Сибири, совсем не изучены Арктика и Крайний Север азиатской части страны. Необходимо отметить и то, что существует прямая зависимость между количеством зарегистрированных в отдельных регионах видов макромицетов и продолжительностью микологических исследований, а также числом микологов, проводящих работы в конкретных районах, областях, республиках. По литературным сведениям, максимальное количество макромицетов – в Приморском крае (802 вида) и в Эстонии (736 видов), наименьшее – на Таймырском полуострове (46 видов), в Туркменской (165) и Казахской ССР (188). Для остальных районов среднее число видов колеблется от 300 до 500. Видовой состав и возраст древостоя, разнообразие типов леса, лесистость и географическое положение района исследований, безусловно, влияют на количество макромицетов и на соотношение экологических групп грибов, однако везде, кроме Крайнего Севера и Юга, на первом месте стоят сапротрофы, а на втором – микоризообразователи, т. е. симбиотрофы. В Таймырских тундрах симбиотрофы составляют 78,5% от всего количества зарегистрированных видов. Учитывая наличие здесь вечной мерзлоты, заболоченности и низкой аэрации болотно‑глеевых почв, крайне неблагоприятных для растений гидротермических условий обитания и широко развитой у кустарниковых форм ив и берез поверхностной корневой системы, можно предположить почти полную микотрофность тундровых растений. Большинство грибов образуют микоризу на корнях карликовой березы, стелящейся по земле, их плодовые тела (березовики, осиновики) по высоте часто превосходят своих симбионтов – деревьев. Короткое северное лето корректирует время появления и продолжительность плодоношения грибов, но масса их достигает внушительных размеров. При переходе от зоны тундры к лесотундре количество видов макромицетов возрастает довольно резко. Здесь ярко выражена особенность симбиотрофных макромицетов – они теряют облигатность связей с определенной древесной породой. Верные спутники сосны, сопровождающие ее на всей территории распространения, встречаются в лесотундрах Кольского полуострова без нее, т. е. ареалы грибов шире, чем у симбионтов. Набор видов для различных древесных пород, растущих в неблагоприятных климатических условиях на каменистых и щебнистых маломощных почвах, оказывается удивительно однообразным. Так, грибы, растущие южнее, образуют микоризу только с сосной, пихтой, кедром, приурочены к ели. Ряд характерных для дубрав симбиотрофов вступают во взаимоотношения с березой. Таким образом, в крайне неблагоприятных для деревьев условиях обитания обязательность симбиотических связей между партнерами ослабляется вплоть до полного исчезновения и набор видов, образующих микоризу с различными деревьями, становится универсальным. Растения, находясь в экстремальных условиях, теряют жесткость связей с определенными, верными для них, грибами, вступая в симбиоз с теми, которые в силу экологических особенностей могут обитать в данных условиях. При продвижении от лесотундры к тайге и далее к зоне хвойно‑широколиственных лесов количество симбиотрофных макромицетов резко увеличивается, достигая максимума в смешанных лесах. В этом же направлении изменяются почвенно‑грунтовые и гидротермические условия обитания для растений, становясь все более оптимальными, растет видовое разнообразие деревьев и кустарников – потенциальных симбионтов макромицетов. Увеличение количества видов микоризообразователей сопровождается облигатностью связей в отношении симбионта, верностью определенной древесной породе, упорядоченным размещением плодовых тел грибов, уменьшением численности каждого вида и относительно быстрой сменой слоев. Здесь особенно ярко проявляются конкурентные взаимоотношения между видами, принадлежащими к одной экологической группе. Микологических данных для зоны широколиственных лесов недостаточно, чтобы провести объективный анализ распределения симбиотрофных грибов, но по имеющимся сведениям количество их меньшее, чем в хвойно‑широколиственных лесах. Дальнейший спад числа видов микоризообразующих макромицетов наблюдается в лесостепях и степях. Для искусственных, в частности полезащитных, насаждений степей и полупустынь характерны, как и на Крайнем Севере, отсутствие специфичности видового состава грибов, образующих микоризу с различными деревьями, очень бедный видовой состав и высокая численность грибов, способных плодоносить в крайне неблагоприятных гидротермических условиях названных зон. В горных лесах Молдавии, Крыма, Кавказа и Средней Азии с их высотной поясностью прослеживаются все те закономерности, которые наблюдаются в зональных равнинных лесных сообществах европейской части СССР, т. е. количество видов симбиотрофов увеличивается в зависимости от количества древесных пород и условий обитания для них, а верность грибов определенной древесной породе нарастает в сторону экстремальных пределов произрастания симбионтов. Большую часть территории Сибири и Дальнего Востока занимают леса, в которых процент микоризообразующих грибов несколько снижается по сравнению с лесами европейской части СССР за счет увеличения числа видов, относящихся к другим экологическим группам. Наличие разновозрастных лесных сообществ, находящихся в самых различных по почвенно‑грунтовым условиям местообитаниях, большой набор древесных пород в лесах Дальнего Востока и благоприятная гидротермическая обстановка в них вызывают чрезвычайно выраженное видовое разнообразие симбиотрофных грибов. Сапротрофы включают три основные группы грибов – ксилотрофы, подстилочные и гумусовые макромицеты, составляющие в отдельных регионах до 70% от общего количества видов. На долю остальных групп приходится от 2 до 14 %. Ксилотрофы, т. е. дереворазрушающие грибы, в своем распространении связаны с количеством и качеством субстрата (отмершей древесины). Их видовой состав и обилие являются показателями возраста, лесорастительных свойств и состояния древостоя, интенсивности антропогенных или стихийных (пожары) вмешательств. На фоне меньшего, чем у микоризообразующих грибов, видового разнообразия возрастает число видов с более или менее повсеместным распространением, обладающих широкой экологической пластичностью. В равнинных и горных лесах европейской части СССР, издавна подвергавшихся интенсивной хозяйственной деятельности, ксилотрофы составляют обычно около 20% от общего количества макромицетов, причем на границах распространения лесов видовое разнообразие грибов этой группы резко снижается и сопровождается нарушением обязательной приуроченности к определенному виду субстрата, т. е. сменой либо качества субстрата, либо его состояния. В лесах с благоприятными почвенно‑грунтовыми и гидротермическими условиями (хвойно‑широколиственные, широколиственные) количество дереворазрушающих грибов максимально. Леса Сибири и Дальнего Востока в сравнении с европейскими характеризуются резким подъемом числа дереворазрушающих грибов вследствие большого распространения старовозрастных древостоев и наличия огромных пространств, подверженных в различное время пожарам, в совокупности приводящих к невероятному количеству отмершей древесины – субстрата для ксилотрофов. На Дальнем Востоке к этим факторам прибавляются благоприятные гидротермические условия приморского климата, а именно высокая относительная влажность воздуха и отсутствие резких перепадов температуры. В связи с этим здесь наблюдается максимальное видовое разнообразие ксилотрофов и самый высокий процент этих грибов в экологической структуре группировок макромицетов. Одна из самых многовидных и специфичных для определенного района группа грибов – подстилочные сапротрофы, которые недостаточно изучены и прежде всего из‑за недолговременности существования плодовых тел грибов и их небольших размеров. Как и для ксилотрофов, ведущим фактором для подстилочных сапротрофов является биохимический состав субстрата – лесной подстилки – и его количество. Но если эти параметры для ксилотрофов ограничены относительно небольшими вариациями (около 300 видов произрастающих на территории СССР деревьев и кустарников, 5–6 градаций состояния субстрата), то для подстилочных сапротрофов число вариаций субстрата во много раз выше. Даже в пределах одного участка леса качественный состав, сложение и запасы подстилки различаются очень существенно. На бесчисленное сочетание вариантов субстрата (соотношение опавшей хвои, листьев, веточек, шишек, отмерших частей травяного, мохового, лишайникового покровов) накладывается чрезвычайное разнообразие микроклиматических особенностей местообитания (температура и влажность субстрата), к которым подстилочные сапротрофы очень чувствительны. Поэтому структура группировок подстилочных сапротрофов в сравнении с другими грунтами грибов наиболее сложная. Поскольку запасы субстрата, зависящего от скорости процессов разложения подстилки, подчиняются зональному распространению, видовое разнообразие подстилочных сапротрофов следует ему. Минимальное количество видов и численность подстилочных сапротрофов наблюдаются в однопородных лиственных лесах, где основная роль в разложении органических веществ, заключенных в подстилке, принадлежит микроорганизмам и почвенным животным. В хвойных лесах, характеризующихся высоким процентом трудноразлагаемых веществ, количество подстилочных сапротрофов увеличивается. Максимума видового разнообразия, а часто и обилия подстилочные сапротрофы достигают в хвойно‑широколиственных лесах. Подстилка здесь мощная, разделена на четкие слои и биохимически разнородная. Учитывая это, в зовах и поясах хвойно‑широколиственных лесов европейской части СССР и Дальнего Востока отмечается самый высокий процент подстилочных сапротрофов. В своем распределении гумусовые сапротрофы подчиняются зональным закономерностям гумусообразования. С севера на юг содержание гумуса в почве, а вм
|
||
|
|
Последнее изменение этой страницы: 2021-07-19; просмотров: 126; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.11 (0.018 с.) |