Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Урожаи грибов и их цикличностьСодержание книги Поиск на нашем сайте
Под урожайностью макромицетов в микологии понимают массу видимой их части – спорадически появляющихся плодовых тел грибов, но ведь основная, рабочая часть гриба – мицелий – скрытая от наших глаз в субстрате: лесной подстилке, опаде, древесине, почве. Грибные гифы пронизывают мертвое органическое вещество до уровня внутриклеточных структур, в связи с чем до сих пор не удается достоверно определить запасы мицелия макромицетов ни в лабораторных условиях, ни тем более в природе. Это касается не только грибов, но и многочисленных обитателей лесной подстилки и почвы: водорослей, беспозвоночных животных, микромицетов, бактерий, простейших. Что касается макромицетов, имеются немногочисленные литературные сведения о соотношении массы мицелия и массы плодовых тел, рассчитанные на основе экспериментов, проведенных в лабораторных условиях. Для напочвенных сапротрофов это соотношение составляет 62,6:1, а для симбиотрофов 154:1, т. е. для формирования одного грамма плодового тела гриба в первом случае необходимо 62,6 г мицелия, во втором – 154. Эти цифры внушительны, но сколько же необходимо переработать субстрата, чтобы создать один грамм мицелия? Это пока неизвестно, как и неизвестно физиологическое состояние мицелия в природе, перераспределение вещества и энергии между плодовым телом гриба и мицелием. Исходя из приведенных цифр, мы для ельника Подмосковья (возраст 80 лет, примесь в древостое березы и дуба) рассчитали массу мицелия макромицетов. Она оказалась равной 2,5 т на 1 га и превосходит по величине массу мхов и травянистых растений. Доля мицелия грибов и других подстилочнообитающих организмов по массе составляет 25% от запасов опада и подстилки в этом лесу. Эта цифра может быть и выше, а это значит, что в лесу мы буквально ходим «по живому», не подозревая, какой удивительный мир организмов у нас под ногами скрыт от наших глаз опавшими листьями, травой, мхами! Для конкретного леса основную массу плодовых тел формируют симбиотрофы и ксилотрофы, обладающие крупными плодовыми телами, хотя по количеству видов они могут и уступать напочвенным сапротрофам. Колебания урожаев в одном типе леса могут достигать огромных величин в многолетней динамике в основном за счет периодичности массового плодоношения микоризообразователей и дереворазрушающих грибов. Причины периодичности урожаев съедобных грибов обычно связывают с количеством осадков и температурой конкретного года. Имеются даже попытки прогнозировать урожайность грибов, но поскольку прогноз погоды у нас еще не совсем объективен, предсказание урожаев грибов не состоятельно. При исследовании зависимости урожаев грибов от гидротермических условий учитывают количество осадков, температуру воздуха и почв, предшествующих или совпадающих по времени с их интенсивностью. Однако при этом совершенно не учитываются физиологические циклы развития мицелия, которые наверняка играют ничуть не меньшую роль, чем погодные условия. По нашим наблюдениям, малосходные погодные условия вызывали массовое появление в лесах Подмосковья веселок, ежовика коралловидного с периодичностью соответственно 8 и 5 лет. Речь идет именно о массовом их появлении, так как отдельные экземпляры этих видов изредка встречались и в годы между циклами. Игнорирование физиологической цикличности плодоношения грибов вносит бесконечные разногласия в вопросе о ведущих факторах, определяющих урожайность грибов, а зачастую приводит и к необоснованным суждениям о редкости видов. Физиологическая цикличность урожаев грибов в чистом виде должна проявляться только в годы, приближающиеся по количеству тепла и влаги к среднемноголетнему, в достаточной степени абстрактному вегетационному периоду. Существенную поправку в этот ритм, безусловно, вносят погодные условия конкретного года, давая толчок к массовому плодоношению либо более сухолюбивых, либо, наоборот, влаголюбивых видов. Чтобы разобраться в вопросе, что же определяет массовое появление грибов – погодные условия или физиологическая цикличность, или сочетание обоих фактов, рассмотрим ритмичность плодоношения опенка настоящего или осеннего – одного из самых распространенных и уважаемых грибниками представителей макромицетов. Наблюдения проводились в лесах Подмосковья, но прежде чем переходить к исследованию закономерностей массового появления опенка, необходимо остановиться на некоторых биологических особенностях самого гриба. Осенний опенок космополитный вид, произрастающий на древесине более 200 видов растений, в основном деревьев и кустарников. На территории СССР он встречается в различных природных зонах на видах хвойных и лиственных пород – ели, сосны, пихты, лиственницы, березы, осины, дуба, липы, клена, ясеня, каштана, яблони, сливы, сирени, а также на кустарниках – малине, крыжовнике, смородине. В качестве источника пищи опенок использует как представитель факультативных (необязательных) паразитов углеводы растительных тканей, в большинстве случаев камбия. Оптимальные условия для распространения мицелия и ризоморфов (собранные в темноокрашенные шнуры гифы гриба различной толщины и до нескольких метров длины), а также обилие плодовых тел опенка связывают с влажностью почв и концентрацией кислорода в них. Чем выше содержание кислорода и соответственно ниже содержание углекислого газа, тем выше границы распространения ризоморфов в почве. Во влажных местообитаниях основная масса ризоморфов сосредоточена в верхнем 10‑сантиметровом слое почвы. Развитие мицелия в субстрате зависит также от содержания и соотношения в нем углерода и азота. Причем повышенное содержание азота способствует развитию ризоморфов и стимулирует паразитарную активность опенка, а возрастание количества углерода вызывает активное разрастание воздушного мицелия, определяя сапротрофный образ жизни гриба. В природных условиях опенок может быть и микоризообразователем (у орхидных), и сапротрофом, разлагающим только мертвую древесину, и опасным паразитом, уничтожающим целые массивы лесов. Вопрос о принадлежности опенка осеннего к группе сапротрофных или паразитных дереворазрушающих грибов дискутируется издавна, но большинство советских микологов и лесоводов относят его к факультативным паразитам. Скорее всего, это так, потому что паразитизм опенка проявляется только в определенных, неблагоприятных для деревьев и кустарников условиях, когда они находятся в ослабленном жизненном состояния (высокий уровень корневой конкуренции между деревьями за питательные элементы, влагу, свет, различного рода повреждения корней и стволов). Мы изучали распределение и урожайность опенка осеннего в смешанных (елово‑лиственных) лесах, осинниках и березняках, искусственных насаждениях ели и сосны различного возраста, вырубках Подмосковья в течение нескольких лет. Опенок в эти годы встречался во всех лесах выше 30‑летнего возраста и на вырубках различного возраста. Максимальное количество плодовых тел гриба отмечено во все годы на пнях и валежной древесине ели и березы, на сухостойных деревьях ели, березы, дуба, ивы, лещины (лесной орех). Изредка в отдельные, особенно благоприятные и урожайные, годы отмечалось нахождение опенка на пнях и сухостое осины, черемухи, сосны, сирени. На древесине липы и клена плодовых тел гриба не найдено. В качестве паразита опенок в названных лесах встречался в основном на корнях и у основания стволов ели в искусственных ее посадках в возрасте от 40 до 80 лет. Большое количество плодовых тел сплошь покрывает нижние части стволов деревьев, от уровня земли до высоты одного метра. Это темные леса с почти полным отсутствием напочвенного травяного покрова. Стоящие в рядах деревья (расстояние между рядами около 5 м, между деревьями в рядах – 3 м) к указанному возрасту оказываются в условиях жесткой конкуренции, следствием чего является угнетенное, ослабленное состояние отдельных деревьев. В тканях таких деревьев происходят и физиологические изменения: снижается активность окислительно‑восстановительных процессов, уменьшается количество аминокислот и глюкозидов в корнях. На корнях и у основания деревьев, находящихся в более благоприятных условиях (опушки посадок, зоны их контакта с просеками, лесными дорогами), опят не обнаружено. Механическое же нарушение корневой системы деревьев (обрубка корней) моментально (на первый, второй год) вызывает массовое появление опенка в зоне повреждения. Такая быстрая реакция подтверждена экспериментами с выращиванием опенка в культуре (лабораторные условия), когда на мицелии, выращенном из спор, плодовые тела образовывались через 2–5 мес на древесине ели и сосны. Реже опенок встречается на живых деревьях березы и дуба, и то только тогда, когда они находятся на гранях своего жизненного уровня (возраста) или механически повреждены, или повреждены жуками‑короедами. В качестве субстрата в этих случаях гриб использует отмершие части коры. В отдельные годы массового плодоношения опенок встречается на живом подросте ели, березы, дуба, где молодые деревца образуют непроходимые заросли. Таким образом, опенок как факультативный паразит выступает в роли индикатора угнетенного состояния деревьев и способствует выпадению их из древостоя (изреживанию), выполняя роль санитара и биологического регулятора видового состава и численности растений. При выпадении отдельных деревьев образуются окна в древесном пологе, изменяются экологические условия для рядом стоящих деревьев, т. е. увеличивается площадь питания корневой системы, большее количество осадков проникает к поверхности почвы, изменяется освещенность, перестраивается напочвенный покров. Проникновение ультрафиолетовой части солнечной радиации к поверхности корневых лап и оснований стволов деревьев может служить причиной подавления паразитарной активности опенка. Более типичным в названных лесах является сапротрофный тип развития опенка. Он встречается на пнях и валеже под пологом леса и на вырубках. Пни лиственных пород деревьев заселяются грибом обычно в первый же год их существования независимо от того, было дерево поражено паразитной формой опенка или не было. После рубки ели, в случае, если при жизни на ней не было опенка, появление плодовых тел гриба наблюдается не ранее, чем через 2 года и продолжается на протяжении 8 лет. На вырубках и просеках опенок плодоносит в течение 10– 15 лет до почти полного разложения древесины, скорость которого зависит от экологической обстановки конкретного местообитания. Так, на протяжении 1969–1980 гг. наблюдалось наряду с редким и единичным появлением опенка в определенных местообитаниях и на определенном субстрате массовое его плодоношение. Сроки начала плодоношения опенка осеннего в лесах Подмосковья колеблются в интервале 10 дней, с 26 августа по 7 сентября. Исключение составлял только 1972 год, необычайно жаркий и сухой, когда горели торфяники и леса в Московской, Рязанской, Владимирской, Горьковской, Костромской и других областях. В то лето грибов практически не было, температура подстилки в лесах в июле поднималась до невероятной отметки +37° и при полном отсутствии осадков мицелий многих грибов сильно пострадал, а некоторых погиб (напочвенных сапротрофов, обладающих мелкими плодовыми телами). Опенок в этот год появился лишь 18 сентября, после выпадения дождей в очень небольшом количестве. Многие микологи связывают годы массового плодоношения опенка с резкими перепадами температуры воздуха в конце лета и увеличением количества осадков, вызывающих высокие показатели относительной влажности воздуха. Исходя из этого, мы проанализировали погодичное количество осадков в течение вегетационного периода, т. е. изменение влажности субстрата до средних сроков появления плодовых тел опенка (к 1 сентября), количество выпавших осадков подекадно в августе, а также сумму положительных температур, накопленных к 1 сентября, изменения средних, минимальных и максимальных температур воздуха ежемесячно и подекадно.
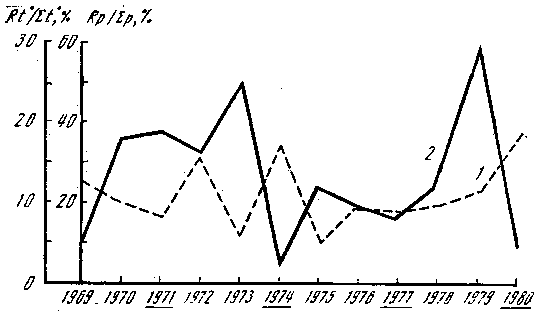
Рис. 8. Количество осадков, суммы положительных температур и плодоношение Armillariella mellea Rt0 – результирующая температура месяцев (май – август); Rp – результирующая осадков тех же месяцев, мм; Σр – сумма осадков, выпавших к 1 сентября; Σt0 – сумма положительных температур, накопленных к 1 сентября; 1 – Rt0/Σt0, %; 2 – Rp/Σр, %. Подчеркнуты годы массового плодоношения опенка
Рассмотрим вначале зависимость урожайности опенка осеннего (Armillariella mellea) от погодичных изменений температуры. На рис. 8 показана величина, отражающая отношение результирующей накопленных положительных температур четырех месяцев (май – август) к сумме температур за эти месяцы. Она указывает на равномерность накопления тепла; чем ближе к 0° С, тем равномернее распределены температуры в течение исследуемого периода. Абсолютная величина результирующей колеблется от 25 до 320° соответственно в 1979 и 1972 гг. Максимальное количество накопленных положительных температур в июне наблюдалось только в 1969 г., в июле – в 1973, 1974, 1975, 1977 и 1979 гг., в августе – в 1970, 1971, 1972, 1976, 1978 и 1980 гг. Как видно из рисунка, накопление температур происходит с выраженными колебаниями минимума и максимума, период колебаний составляет 2 года. На этом же рисунке отмечены годы с массовым плодоношением опенка. Анализ показывает, что периоды колебаний исследуемых явлений не совпадают. Были рассмотрены и показатели накопленных к 1 сентября (началу плодоношения опенка) положительных температур. Во все исследуемые годы она к этому времени равнялась 2 000–2 500°. Различия эти определяются суммой температур март – конец июня. Месячные и подекадные средние, максимальные и минимальные температуры воздуха вариабельны, а их анализ не дает никаких оснований связывать их величину и изменчивость с периодичностью плодоношения опенка. Можно лишь отметить, что массовому появлению плодовых тел гриба соответствуют величины абсолютного минимума температур – от –1 до +3°, абсолютного максимума – от +18 до +22° в августе. Как было отмечено, наиболее резкие различия в накоплении суммы положительных температур наблюдаются в весенние месяцы и особенно в апреле. Амплитуда колебаний этой величины по годам достигает 275°, однако высоким урожаям опенка сопутствуют значения суммы положительных температур в очень узких пределах – 80–90°. Это единственная закономерность, которую нам удалось обнаружить. Количество выпавших осадков в исследуемые годы резко различается в своем распределении по месяцам, декадам и сумме к 1 сентября. Такого четкого и ритмичного колебания минимума и максимума, которое было отмечено для суммы положительных температур, в случае с осадками не наблюдается. Об этом свидетельствуют данные, представленные на рис. 9. Отношение результирующей четырех месяцев (май – август) к сумме осадков, выпавших за эти месяцы, указывает на резко выраженную их неравномерность. Причем никакой зависимости между массовым плодоношением опенка и распределением осадков не наблюдается. Однако можно отметить, что урожайным годам соответствует более или менее равномерное распределение осадков, когда величина отклонения их в различные месяцы не превышает 5–8%. Между абсолютным количеством осадков, выпавших к началу плодоношения опенка, а также ежемесячным и подекадным их режимами и появлением плодовых тел гриба зависимости не найдено. Некоторую информацию о связи этих явлений дает количество осадков, выпавших в последнюю декаду августа, перед непосредственным началом плодоношения. Сумма их колеблется от 3 до 5 в 1971, 1974 и 1977 гг. до 32 мм в 1980 г.
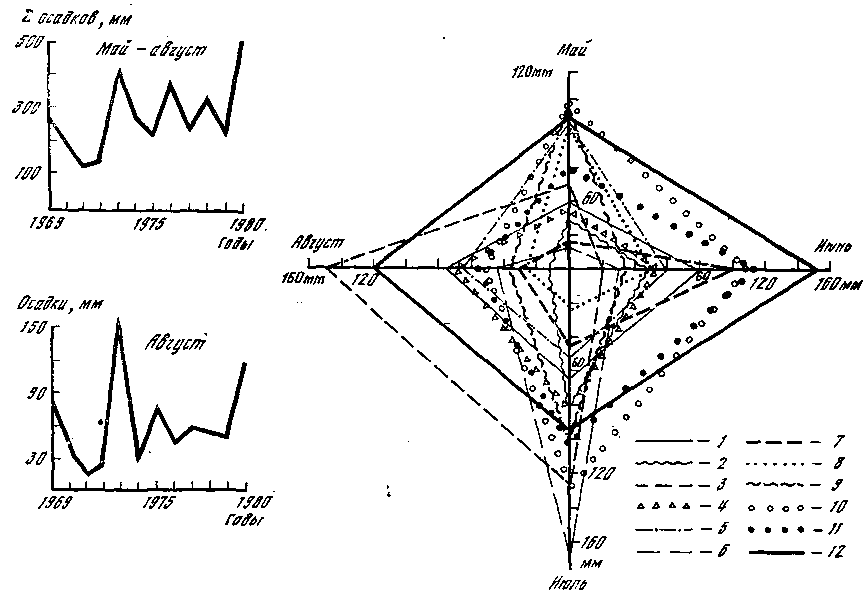
Рис. 9. Амплитуда колебаний выпавших в 1969–1980 гг. осадков 1 – 1969; 2 – 1970; 3 – 1971; 4 – 1972; 5 – 1973; 6 – 1974; 7 – 1975; 8 – 1976; 9 – 1977; 10 – 1978; 11 – 1979; 12 – 1980
Одновременный анализ распределения температур и осадков в связи с цикличностью плодоношения опенка показывает, что урожайные годы характеризуются довольно резкими различиями в показателях суммы накопленных положительных температур и количества выпавших к 1 сентября осадков. В 1971 г. максимальное количество осадков выпало в июле на фоне равномерного накопления температур; 1974 г. (год массового появления опенка) характеризуется жарким июнем при влажном июле; 1977 г. – максимально влажным июнем и жарким июлем и, наконец, 1980 г. – самыми высокими показателями температуры и осадков в августе. Периодичность массового развития опенка, как видно из рис. 9, составляет 3 года и не совпадает с рассмотренными показателями температурного режима и режима влажности. По‑видимому, такая цикличность урожаев опенка связана с иными причинами, такими, как, например, физиологические особенности развития гриба, в частности накопление достаточного количества веществ и энергии в мицелии. Периодичность плодоношения макромицетов зависит от сочетания очень многих факторов, На рис. 10 показана многолетняя динамика количества видов и массы грибов различных экологических групп в одном и том же типе леса. Амплитуда колебаний массы грибов всех групп равна 14,8 кг/га, т. е. приближается к среднемноголетним значениям урожая плодовых тел. Такие различия определяются экстремальными годами, каковым оказался 1972 год. В основном же для лесов Подмосковья различия в урожайности грибов составляют 5–6 кг/га и совпадают по цикличности с суммой накопленных положительных температур за период май – сентябрь. Величина этого цикла – 2 года, а поскольку основу массы грибов составляют макромицеты с крупными плодовыми телами, относящиеся к симбиотрофам, можно считать, что периодичность урожаев таких грибов, как сыроежки, грузди, рядовки, паутинники, белые и березовики, составляют именно 2 года.
Рис. 10. Динамика трофической структуры и биомассы макромицетов (липо‑ельник зеленомошно‑волосистоосоковый) Число видов: 1 – симбиотрофов; 2 – ксилотрофов; 3 – гумусовых сапротрофов; 4 – подстилочных сапротрофов; 5 – сапротрофов на опаде; 6 – грибов других трофических групп. Жирной линией обозначена масса макромицетов, кг/га (абс. сухой вес); N – число видов; М – масса грибов
Наибольшей стабильностью в отношении видового состава, численности и массы обладают ксилотрофы и гумусовые сапротрофы, мицелий которых не испытывает в силу своего пространственного положения (защищенности) таких резких колебаний температуры и влажности, как мицелий грибов, расположенных в верхних слоях подстилки. Кроме того, как указывалось ранее, ксилотрофы имеют специфические приспособления для перенесения неблагоприятных погодных условий. Однако самыми консервативными в отношении количества видов и массы оказываются в многолетней динамике сапротрофы на опаде. Объясняется это уникальными физиологическими особенностями плодовых тел этих грибов. На опавших хвоинках и веточках хвойных пород постоянно присутствует в «увядшем» состоянии большое количество хрящеватых тонких (до одного миллиметра) ножек плодовых тел с зачатками шляпок (рис. 10). При повышении относительной влажности приземного слоя воздуха до 70–80% они тут же восстанавливают тургор тканей с минимальной затратой на этот процесс веществ и энергии. Подстилочные сапротрофы наиболее уязвимы в отношении погодных условий, так как их мицелий, находящийся в различных слоях подстилки, слабо защищен от перепадов температуры и влажности, у них нет симбионтов, которые могли бы помочь им в защите, и нет специальных приспособлений, как у ксилотрофов (ризоморфов), для перенесения неблагоприятных условий. Масса и количество видов макромицетов в течение одного сезона постепенно увеличиваются с мая, достигая максимальных значений в августе – начале сентября, а затем резко или медленно в зависимости от погодных условий конкретного года уменьшаются. Заканчивается плодоношение всех представителей экологических групп грибов в октябре – начале ноября. Самыми холодостойкими являются некоторые виды ксилотрофов и симбиотрофов, замерзающих на корню (опята, белые, рядовки). Такое распределение массы макромицетов характерно для лесов умеренного климата. В районах с резко выраженным недостатком тепла за вегетационный период (на севере) и влаги (на юге) сроки роста, увеличения размеров и массы плодовых тел грибов укорачиваются по сравнению, например, с Подмосковьем в 2–3 раза. Реакция мицелия грибов на дефицитные тепло и влагу моментальная, почти как у сапротрофов на опаде. Здесь не каждый год создаются благоприятные погодные условия для образования плодовых тел грибов, но, когда это происходит, масса их огромна – 80–100 кг/га, а количество грибов на одном квадратном метре достигает 10 экз. Обычно каждый гриб появляется в определенное для него время, однако в некоторые годы чисто погодные условия могут сдвигать сроки плодоношения. Так, например, в лесах Подмосковья понижение дневной температуры в июле 1976 г. до +10–12°, сопровождавшееся затяжными моросящими дождями, спровоцировало появление плодовых тел типично осенних видов – волнушки розовой и черного груздя. Отклонения в температуре и количестве осадков летом 1983 г. изменили все сроки развития грибов. Жаркая и влажная погода в июне – июле вызвала массовое развитие «колосовиков» – белых и березовиков и затянула их плодоношение на 1,5–2 мес. в отличие от среднемноголетнего (1,5–2 нед). Наступившая за этим жаркая и сухая погода августа исключила плодоношение всех симбиотрофов, связанных по срокам с этим месяцем. Таким образом, на цикличность, величину урожаев, количество видов грибов влияет много факторов, среди которых основными являются физиологическое состояние мицелия и сочетание температуры и влаги в субстрате. Но пи в коем случае нельзя связывать урожайность макромицетов только с одним из них, например с осадками или температурой, как это зачастую у нас делается даже в научной (микологической) литературе.
Лес – среда обитания грибов
Когда мы произносим слово «гриб», перед нашим мысленным взором сразу возникают леса: светлые березняки и сосняки, темные мрачные ельники, влажные и сухие, травяные, моховые, лишайниковые – словом, самые разные. И эта аналогия не случайна, ведь большинство грибов макромицетов (95%) – обитатели лесов. Взаимосвязи грибов с лесом разнообразны и сложны. Они осуществляются через атмосферу, растения, почву, животных, микроорганизмы и могут быть положительными или отрицательными, специфичными или универсальными, контактными или опосредованными. Грибы – полноправные жители лесов, и существовать вне леса они не могут. Для макромицетов в первую очередь важен состав древесных пород в лесу и почвенно‑грунтовые условия, в которых они находятся. Просто ельник или сосняк еще ничего не говорят грибнику о том, сколько и каких грибов он может в них собрать. А это необходимо знать, чтобы не уйти из леса пустым. Охарактеризуем основные типы лесов на территории СССР по признаку произрастания в них различных видов грибов. Одной из самых распространенных древесных пород считается ель, образующая чистые и смешанные насаждения в смеси с березой, осиной, ольхой, сосной, пихтой, кедром и лиственницей. Ельники обычно занимают относительно богатые суглинистые и супесчаные почвы и редко бывают однопородными. В зависимости от почвенно‑грунтовой влагообеспеченности и по мере ее увеличения ельники образуют ряд типов: неморальные (в основном папоротниковые), кислично‑черничные и черничные, долгомошные и сфагновые, болотно‑травяные. Ельники не очень богаты съедобными грибами. Здесь можно найти: рыжики (больше всего их в молодых мертвопокровных густых ельниках); белые (еловая, светлоголовая форма); грузди: желтый, обыкновенный или гладыш (во влажных и сырых ельниках); сыроежки: буреющую (бордово‑коричневая шляпка, мякоть ножки и шляпки при разламывании сереет и имеет селедочный запах); Келе (шляпка фиолетово‑пурпурно‑бурая, ножка красновато‑фиолетовая), неприятную (шляпка бледно‑красновато‑сероватая, ножка белая), болотную (шляпка красно‑буроватая, ножка белая или розоватая, растет в долгомошных и сфагновых ельниках). Кроме названных грибов, в ельниках растут и малоизвестные съедобные грибы, такие, например, как мокруха еловая, шампиньон лесной, говорушка бокаловидная. Из несъедобных и ядовитых в ельниках можно встретить иногда в большом количестве паутинник вонючий, мухомор порфировый (шляпка фиолетово‑пурпурно‑серая с крупными грязно‑белыми лоскутами), ложноопенок серно‑желтый. В отдельные годы в различных ельниках можно много собрать осенних опят, свинушек. Сосновые леса в отличие от еловых занимают бедные песчаные, каменистые, торфяные почвы. По отношению к почвенной влагообеспеченности они, как и ельники, образуют ряды. Самые сухие сосняки – лишайниковые и вересковые, за ними (по мере увеличения влажности почв) идут зеленомошные, брусничные, черничные, долгомошные, сфагновые, и, наконец, самые «болотные» – пушициево‑ и осоково‑сфагновые. По сравнению с ельниками сосняки очень богаты грибами, как съедобными, так и ядовитыми. С ранней весны, как только сойдет снег, появляются в сосновых лесах строчки, которые после соответствующей обработки (прокипятить не менее 20 мин и отвар обязательно слить) вполне можно употреблять в пищу. С июня по опушкам сосняков и в молодняках «высыпают» плодовые тела масленка зернистого (шляпка желто‑охристо‑коричневая, ножка без кольца), а в августе к нему присоединяется масленок поздний (шляпка густослизистая шоколадно‑темно‑коричневая, ножка с кольцом). Они образуют целые поля, только жаль, что большая часть маслят бывает червивая. В конце июля – начале августа в сосняках грибное изобилие. Группами попадаются темноголовые боровики, крепкие, красные рыжики (в отличие от хрупких зеленоватых еловых). В зеленомошных сосняках особенно много моховиков желто‑бурых (шляпка грязно‑желтая, ножка желтоватая, мякоть на изломе быстро синеет, трубчатый слой очень плотный, табачно‑бурого цвета). К тому же роду, что и моховик, относится решетник, или козляк, тоже очень распространенный в сосняках гриб (шляпка неопушенная, блестящая, оранжево‑коричневая, мякоть плотная, желтовато‑буроватая, трубчатый гименофор с широкими неправильно‑угловатыми порами). Из млечников чаще всего в сосняках встречается горькушка (шляпка сухая, темно‑красно‑бурая с сосочком в центре, млечный сок белый, обильный и едкий), млечник серо‑розовый (шляпка сухая, серовато‑розоватая, млечный сок водянисто‑белый, не едкий, мякоть с сильным запахом кумарина) и млечник белый (шляпка желтовато‑белая, млечный сок водянисто‑белый, не едкий, пластинки при надавливании становятся оливково‑охристыми). Сыроежек в сосновых лесах тоже довольно много. Одна из самых распространенных – подгруздок черный, обычно грибниками непричисляемый к сыроежкам. Внешне гриб напоминает грузди, но у него нет млечного сока. Растет в основном в зеленомошных сосняках и образует большие скопления плодовых тел. Шляпка довольно больших размеров (до 15 см), ножка короткая (3–4 см) и толстая, кожица на шляпке гладкая, липкая, грязно‑бурая. Мякоть плотная, на изломе быстро становится розовато‑серого, а затем черного цвета. Это хороший съедобный гриб, но, как и маслята, очень часто бывает червивым. В болотистых сосняках, особенно по краям, встречается крупная оранжевожелтая сыроежка. Мякоть шляпки и ножки серая, за что она и получила название сыроежки сереющей. Вместо с ней попадается сыроежка болотная с красно‑буроватой, иногда пятнистой шляпкой и белой или розоватой ножкой. В сосняках раздолье для съедобных рядовок. Самая знаменитая из них зеленушка, приравненная по вкусовым качествам к грибам первой категории. Шляпка у нее слабослизистая или клейкая оливково‑буроватая, а по краям зеленоватая. Похожа на зеленушку и другая рядовка – серая, но шляпка у нее оливково‑серо‑бурая и пластинки более ярко окрашены (с лимонно‑желтым оттенком). Встречается в те же сроки (август–октябрь), что и зеленушка, и также обильна. К двум названным съедобным рядовкам относится и рядовка чешуйчатая, приуроченная к зеленомошным соснякам и образующая здесь в августе‑сентябре большие скопления плодовых тел группового распределения. Шляпка у нее радиально‑чешуйчатоволокнистая, коричнево‑бурая, а ножка довольно высокая (5–10 см) и тонкая (1–1,5 см). Близка к ней по окраске шляпки и тоже съедобна бело‑бурая рядовка, во отличается мучным запахом мякоти и короткой ножкой (3–5 см), а также отсутствием чешуек и слабой слизистости на шляпке. К малоизвестным съедобным грибам в сосняках с примесью дуба относится синяк, настораживающий грибников, по‑видимому, ярко‑синим цветом мякоти на изломе. Он относится к трубчатым грибам, шляпка и ножка палевого или бледно‑терракотового цвета. Растет в изобилии с конца июля до сентября обычно вдоль дорог или в разреженных сосняках и суборях. Во многих районах СССР грибниками игнорируется и другой, очень распространенный, особенно в зеленомошных сосняках, съедобный гриб‑колпак кольчатый. По внешнему виду он напоминает известный всем валуй, но шляпка у него сухая, палево‑ или глинисто‑желтая, на верхушке с мучнистым налетом, а пластинки широкие, охристо‑ржаво‑бурые, на ножке имеется пленчатое кольцо, как у шампиньона. Это крупный гриб (шляпка диаметром до 8 см, ножка высотой 8–10 см), образующий с середины лета до конца осени скопления плодовых тел, четко приуроченных к сплошному зеленому ковру мхов. Много в сосняках ядовитых и несъедобных грибов с крупными плодовыми телами. Из несъедобных в первую очередь надо отметить желчный гриб, растущий на почве или древесине. В последнем случае обычно в переувлажненных сосняках. Он очень похож на березовик или белый со светлоокрашенной шляпкой по форме и окраске, но отличается от них очень горькой и розовеющей на изломе мякотью, а также темно‑бурым сетчатым рисунком на ножке. Гриб не смертельно ядовит, но может вызывать поражение печени. К условно съедобным относится свинушка толстая, обитающая на пнях сосны или около них. Отличается от свинушки тонкой войлочно‑бархатистой, черновато‑бурой, толстой ножкой и желтоватой, горькой мякотью. При большом количестве отличных съедобных грибов, растущих в сосняках, лучше ее не собирать. Из мухоморов в сосновых лесах встречаются красный, желтый, оба ядовиты. Последний приурочен к зеленомошным соснякам, имеет относительно небольшую шляпку (диаметр до 7 см) желтого цвета, рубчатую по краю, с очень редкими белыми лоскутками. Ножка тонкая, белая, с небольшим вздутием у основания. Ядовитым считается и ложнодождевик обыкновенный, который встречается повсеместно в различных сосняках, иногда в большом количестве, особенно вдоль дорог. Он представляет собой шар диаметром до 10 см и более палево‑желто‑охристого цвета, покрытый крупными чешуйками. Внутренность шара черно‑фиолетового цвета, на изломе напоминает черную икру. Березовые леса, как и сосновые, распространены на огромных пространствах. Кроме того, береза в качестве примеси присутствует почти во всех хвойных лесах, чем значительно обогащает в них видовое разнообразие грибов. Березняки по праву считаются грибными, так как с различными видами берез связано плодоношение большого количества ценных съедобных грибов. Из них на первом месте стоит, конечно, белый (березовая форма) с очень светлой беловато‑буроватой, иногда охристой шляпкой и более тонкой, чем у боровиков, ножкой. Очень разнообразны в названных лесах подберезовики, резко отличные по форме и окраске. В болотных березняках встречается белая форма березовика. В молодости шляпка чисто‑белая, к старости становится грязно‑белой с зеленоватым оттенком, ножка довольно тонкая и длинная. На одной болотной кочке иногда умещается до полусотни утопающих во мху грибов. В более сухих березняках можно встретить березовик окисляющийся с желто‑бурой шляпкой и розовеющей на изломе мякотью. Ножка у него толще, чем у болотного, и покрыта черно‑бурыми чешуйками. Бывают березовики с почти черными шляпками, толстыми ножками, покрытыми черно‑бурыми чешуйками. Они обычно растут в сыроватых березняках и по краям сфагновых болот. В сухих березовых лесах с мая появляется березовик обыкновенный. Шляпка у него достигает диаметра до 20 см, серовато‑буроватая, ножка длинная, нередко расширяющаяся к основанию, покрытая почти черными чешуйками. Нередко встречается в березняках и осиновик желто‑бурый. Он отличается от истинного подосиновика очень плотной желто‑бурой шляпкой, достигающей в диаметре 30 см, белой ножкой с черными чешуйками и мякотью, которая на изломе сначала приобретает розово‑лиловый цвет, а затем грязно‑серый. С березой связаны и верны ей многие виды млечников. Среди них особое место принадлежит груздю настоящему. Излюбленное место для него, как и для подосиновика желто‑бурого,– березники на супесчаных и песчаных почвах. Здесь он встречается в большом количестве. Шляпка у груздя очень слизистая с густоопушенным подвернутым вниз краем (до 20 см в диаметре), белая или чуть желтоватая. Млечный сок белый, едкий, на воздухе становится серо‑желтым. Верен березе и черный груздь с такой же слизистой, по краю волосисто‑опушенной, шляпкой зеленовато‑ или оливково‑бурого цвета. В старых березняках часто встречается в большом количестве волнушка, а в молодых – белянка, очень похожая на волнушку, но с белой, опушенной по краю, шляпкой без концентрических зон. Много в березовых лесах сыроежек. С конца июля в разреженных березняках и по опушкам можно видеть «стада» волуев, относящихся к этому роду, и подгруздков белых, очень похожих на грузди, но без млечного сока. Шляпка у подгруздка гладкая, край небахромчатый, часто круто подвернутый книзу. Цвет шляпки грязновато‑желтовато‑белый. Пластинки частые с голубоватым оттенком. К сожалению, как и валуй, часто поражается грибными мухами и комарами. Из других видов сыроежек наиболее распространены в березняках синевато‑зеленая (названа по цвету шляпки) и пищевая с красновато‑буроватой шляпкой и твердой белой ножкой. Одним из самых известных и любимых многими спутников березы можно считать лисичку. С июня на березовых опушках появляются россыпи лисичек. Здесь они «толстые», с завернутым краем палевой шляпки. В глубине леса лисички более ярко окрашены, «тоньше», с почти воронковидной шляпкой. Скорее всего, это разные экологические формы одного вида. Из мухоморов особую любовь к березе питают красный (ядовитый) и серо‑розовый (съедобный). Шляпка у последнего серовато‑розоватая, покрыта многочисленными грязно‑розовато‑белыми лоскутками и бородавками. Ножка почти одноцветна со шляпкой, краснеющая. Большие скопления плодовых тел гриба приурочены обычно к опушкам. Осиновые леса довольно широко распространены на территории СССР. Произрастают они, как правило, на более или менее плодородных почвах и потому, вероятно, не имеют такого количества симбиотрофов. Как и тополевые леса, осинники бедны съедобными грибами (тополь и осина относятся к одному ботаническому роду). Безусловно, верным осине считается осиновик красно‑бурый, отличающийся от желто‑бурового, березового менее плотной мякотью, меньшими размерами шляпки, окрашенной в яркий оранжевый цвет, и мякотью, которая на изломе сначала лиловеет, затем чернеет. Чешуйки на ножке вначале белые, потом темно‑бурые. Особенно много подосиновика в молодых мертвопокровных осинниках с неустойчивым избыточным почвенным увлажнением. Другой, не менее верный спутник осины – груздь осиновый, часто встречающийся и в тополевых лесах. Шляпка у него слизистая, белая с размытыми розовато‑красноватыми пятнами и слабозаметными водянистыми концентрическими зонами. Край шляпки завернут вниз. Млечный сок белый, едкий. Пластинки частые, тонкие, кремово‑розоватые. Ножка короткая, одноцветная, со шляпкой. Из других съедобных грибов в осинниках растут опенок осенний и другой представитель дереворазруша<
|
||||
|
|
Последнее изменение этой страницы: 2021-07-19; просмотров: 70; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.139.104.140 (0.017 с.) |