Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Общие морфологические характеристики эпителиев и образующих их клетокСодержание книги
Поиск на нашем сайте Общие морфологические признаки эпителиев включают: 1) расположение клеток (эпителиоцитов) сомкнутыми пластами, которые образуют плоскостные выстилки, сворачиваются в трубочки или формируют пузырьки (фолликулы); данная особенность эпителиев обусловливается признаками (2) и (3); 2) минимальное количество межклеточного вещества, узкие межклеточные пространства; 3) наличие развитых межклеточных соединений, которые обусловливают прочную связь эпителиоцитов друг с другом в едином пласте; 4) пограничное положение (обычно между тканями внутренней среды и внешней средой); 5) полярность клеток - как следствие признака (4). В эпителиоцитах различают апикальный полюс (от греч. apex - верхушка), свободный, направленный во внешнюю среду, и базальный полюс, обращенный к тканям внутренней среды и связанный с базальной мембраной (см. ниже). Многослойным эпителиям свойственна вертикальная анизоморфия (от греч. an - отрицание, iso - одинаковый, morphe - форма) - неодинаковые морфологические свойства клеток различных слоев эпителиального пласта; 6) расположение на базальной мембране - особом структурном образовании, которое находится между эпителиеми подлежащей рыхлой волокнистой соединительной тканью; 7) отсутствие сосудов; питание эпителия осуществляется путем диффузии веществ через базальную мембрану из сосудов соединительной ткани. Различное удаление отдельных слоев многослойных эпителиев от источника питания, вероятно, усиливает (или поддерживает) их вертикальную анизоморфию; 8) высокая способность к регенерации - физиологической и репаративной - осуществляется благодаря камбию (включающему стволовые и полустволовые клетки) и обусловлена пограничным положением эпителиев (определяющим значительную потребность в активном обновлении быстро изнашивающихся эпителиоцитов). Камбиальные элементы в одних эпителиях сконцентрированы в их определенных участках (локализованный камбий), в других - равномерно распределены среди остальных клеток (диффузный камбий). Взаимодействие эпителия с другими тканями проявляется как в процессе внутриутробного развития, так и после рождения. Основной тканью, с которой эпителий осуществляет постоянные индукционные взаимодействия, является связанная с ним рыхлая волокнистая соединительная ткань, которая не только обеспечивает питание эпителия за счет имеющихся в ней сосудов, но оказывает на него регуляторные влияния. Изменение нормальных взаимоотношений указанных тканей может вызывать нарушение их роста и дифференцировки (в частности, явиться механизмом развития опухолей, например, молочной и предстательной желез). Неэпителиальные клетки в пласте эпителия. В пласте эпителия среди его клеток всегда располагаются отдельные неэпителиальные клетки, взаимодействующие с эпителиоцитами. Наиболее многочисленными из них являются внутриэпителиальные лимфоциты, реже обнаруживаются другие лейкоциты. В некоторых эпителиях в значительном количестве содержатся отростчатые клетки нескольких видов, имеющие неодинаковое происхождение и выполняющие разные функции - пигментные клетки (меланоциты), дендритные антиген-представляющие клетки, а также клетки Меркеля (тактильные эпителиоциты). Эпителии как источник опухолей человека. Эпителии часто служат источником развития опухолей человека, в частности, наиболее распространенных злокачественных новообразований - раков. Этому, вероятно, способствует свойственная этим тканям высокая активность процессов регенерации, при которой могут возникать или усиливаться повреждения генетического аппарата клеток. Злокачественные опухоли способны развиваться из покровного и железистого эпителиев; в последнем случае они называются аденокарциномами (от греч. adenos -железа и karkinos - рак). В высокодифференцированных раках в большей или меньшей степени сохраняются морфологические особенности (цитоархитектоника, цитологические характеристики) и функциональные признаки (выработка слизи, ферментов, гормонов, экспрессия специфических цитокератинов), свойственные тем или иным эпителиям. В малодифференцированных опухолях они утрачиваются, и эпителиальную природу опухоли удается установить лишь путем иммуноцитохимического выявления цитокератинов (см. ниже) в ее клетках. Морфологические особенности эпителиоцитов варьируют в широких пределах, различаясь как в разных эпителиальных тканях, так и между отдельными клетками в пределах одной ткани. Эти особенности тесно связаны с функцией клеток и их положением в эпителиальном пласте. Форма эпителиоцитов служит важным классификационным признаком как отдельных клеток, так и эпителиальных пластов в целом, Выделяют плоские, кубические и призматические (столбчатые, или цилиндрические) клетки. Эпителиоцитам, как уже отмечено выше, свойственна полярность. Ядро эпителиоцитов может иметь различную форму, которая обычно соответствует форме клетки: в плоских клетках оно дисковидное, в кубических - сферическое, в цилиндрических - эллипсоидное. В большинстве клеток ядро сравнительно светлое (преобладает эухроматин), содержит хорошо заметное крупное ядрышко, однако в ороговевающих эпителиях по мере дифференцировки клеток оно уменьшается, уплотняется, распадается и лизируется - подвергается кариопикнозу, кариорексису и кариолизису. Цитоплазма эпителиоцитов содержит все органеллы общего - значения, а в некоторых клетках - также органеллы специального значения, обеспечивающие функции данных клеток. В клетках железистого эпителия хорошо развит синтетический аппарат. В связи с полярностью клеток органеллы распределены в их цитоплазме неравномерно. Цитоскелет эпителиоцитов хорошо развит и представлен микротрубочками, микрофиламентами и промежуточными филаментами. Последние в эпителиоцитах особенно многочисленны и называются тонофиламентами. При фиксации, склеиваясь друг с другом, они могут образовывать крупные агрегаты, выявляемые под световым микроскопом и описанные под названием тонофибрилл. Цитокератины - белки, образующие тонофиламенты, которые специфичны для клеток эпителиальных тканей. Исключение составляет эндотелий, для которого характерны виментиновые промежуточные филаменты. Идентифицировано около 30 различных форм цитокератинов, Которые представлены двумя типами: кислыми кератинами (тип I) и основными кератинами (тип II). Выработка каждого вида цитокератина кодируется особым геном. Тонофиламенты образованы гетерополимерным комплексом, включающим не менее двух различных цитокератинов (кислый + основной). Для конкретного вида эпителия (а в многослойных эпителиях - для каждого слоя) характерен определенный набор цитокератинов, экспрессию которых рассматривают как маркер дифференцировки эпителиальных клеток. Изменения нормальной экспрессии цитокератинов могут указывать на нарушения дифференцировки клеток и в ряде случаев служить важным диагностическим признаком их злокачественного перерождения. При ряде заболеваний кожи и слизистых оболочек в эпителиоцитах обнаружены изменения кератинов, связанные с мутациями соответствующих генов. Поверхности эпителиоцита (латеральная, базальная, апикальная) обладают отчетливой структурно-функциональной специализацией, которая особенно хорошо выявляется в однослойном эпителии.
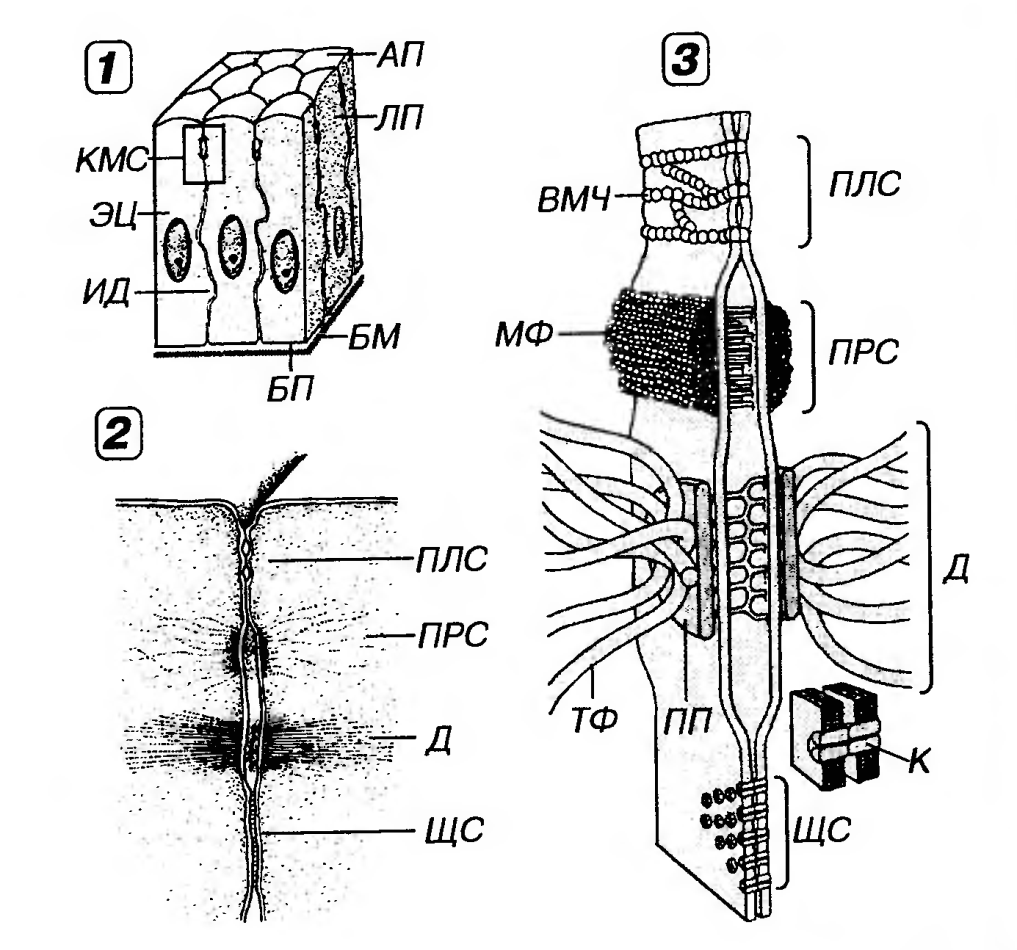
Рис. 1. Межклеточные соединения эпителиоцитов в области их латеральной поверхности. 1 - область расположения комплекса межклеточных соединений (выделена рамкой), 2 - вид межклеточных соединений на ультратонких срезах, 3 - трехмерная схема строения межклеточных соединений (по К. Де Дюву, 1987, с изменениями). БМ - базальная мембрана, БП - базальная поверхность, АГ) - апикальная поверхность, ЛП - латеральная поверхность эпителиоцитов, KMC - комплекс межклеточных соединений, ПЛС - плотное соединение, ПРС - промежуточное соединение, Д - десмосома, ИД - интердигитации, ВМЧ - внутримембранные частицы, ПП - пластинка прикрепления, МФ - микрофиламенты. ТФ - тонофиламенты, ЩС - щелевое соединение, К - коннексоны. Латеральная поверхность эпителиоцитов обеспечивает связь клеток друг с другом за счет специализированных участков - межклеточных соединений, или контактов (рис. 1). Благодаря последним эпителиоциты формируют пласты, что служит важнейшим отличительным свойством организации эпителиальных тканей. Межклеточные соединения подразделяются на два основных вида: 1. Механические соединения - обусловливают механическую связь эпителиоцитов друг с другом. В их число входят плотные соединения, промежуточные соединения, десмосомы, интердигитации; 2. Коммуникационные соединения - (от лат. communicatio - сообщение) обеспечивают химическую (метаболическую, ионную и электрическую) связь между эпителиоцитами. К ним относятся щелевые соединения. (1) Плотное соединение (zonula occludens - поясок замыкания) - наиболее тесный контакт клеток из всех известных в природе. Представляет собой область частичного слияния наружных листков плазмолемм двух соседних клеток (см. рис. 1), которая блокирует распространение веществ по межклеточному пространству (обеспечивая тем самым барьерную функцию эпителия и регулируемость транспорта веществ через эпителиальный пласт). Это соединение также препятствует свободному перемещению и смешиванию функционально различных внутримембранных белков, локализующихся в плазмолемме апикальной и базолатеральной поверхностей клетки, что способствует поддержанию ее полярности. Плотное соединение имеет вид пояска шириной 0.1-0.5 мкм, окружающего клетку по периметру (обычно у ее апикального полюса) и состоящего из анастомозирующих тяжей внутримембранных частиц. Эти частицы образованы белком окклюдином; каждая из них представляет собой область точечного слияния плазмолемм двух соседних клеток. Проницаемость плотных соединений тем ниже, чем выше число тяжей таких частиц. Для поддержания целостности этих соединений необходимы двухвалентные катионы (Са2+, Mg2+). Они могут динамично перестраиваться (вследствие изменений экспрессии и степени полимеризации окклюдина) и временно размыкаться (например, при миграции лейкоцитов через межклеточные пространства). (2) Промежуточное соединение, или опоясывающая десмосома onula adherens - поясок сцепления) локализуется на латеральной поверхности эпителиоцита между областью расположения плотного соединения и десмосом (что обусловило его первое название). Охватывает клетку по периметру в виде пояска (см. рис. 1), на сечении имеющего сходство с десмосомой (что послужило основанием для второго названия). В области промежуточного соединения обращенные к цитоплазме листки плазмолеммы утолщены, образуя пластинки прикрепления, которые содержат актин-связывающие белки α-актинин, винкулин и плакоглобин (последний обнаруживается также в десмосомах). К этим пластинкам прикрепляются элементы цитоскелета - актиновые микрофиламенты, вплетающиеся также в терминальную сеть. Межклеточная щель расширена до 15-20 нм и заполнена умеренно плотным веществом, в состав которого входит адгезивный трансмембранный гликопротеин Е-кадгерин, обеспечивающий в присутствии ионов Са2+ связь между соседними клетками. Со стороны цитоплазмы в области промежуточного соединения к Е-кадгерину через α -актинин и винкулин прикрепляются актиновые микрофиламенты, что обусловливает связь цитоскелета с компонентами межклеточного вещества. (3) Десмосома (macula adherens - пятно сцепления) - состоит из утолщенных и уплотненных участков цитоплазматического листка плазмолемм двух соседних клеток - пластинок прикрепления, разделенных межклеточной щелью (см. рис. 1). Пластинки прикрепления имеют дисковидную форму (диаметр около 0.5 мкм, толщина 15 нм) и служат участками прикрепления к плазмолемме промежуточных филаментов (тонофиламентов). Они содержат особые белки - десмоплакины, плакоглобин и десмокальмин. Межклеточная щель в области десмосомы имеет ширину около 25 нм и заполнена материалом низкой электронной плотности, часто поперечно исчерченным и содержащим в центре линейное уплотнение (центральная, или промежуточная линия). В межклеточном материале десмосомы находятся десмоколлины и десмоглеины - трансмембранные Са2+-связывающие адгезивные белки, которые, взаимодействуя с белками пластинок прикрепления, связывают их в единую систему. Десмосомы разбросаны по поверхности клетки; они, как и промежуточные соединения, служат участками, опосредующими связь элементов цитоскелета (внутриклеточного компонента) с компонентами межклеточного вещества. Повреждение десмосом посредством антител к их компонентам служит главным механизмом патогенеза вульгарной пузырчатки - тяжелого, а в прежние годы смертельного аутоиммунного заболевания кожи и слизистых оболочек. Клетки в многослойных эпителиях утрачивают связи друг с другом, округляются, а их тонофиламенты отсоединяются от пластинок прикрепления и образуют скопления вокруг ядра. Внутри эпителия формируются пузыри, которые вскрываются с образованием эрозий. Связь эпителия с подлежащей соединительной тканью, однако, при этом заболевании сохраняется. (4) Интердигитации - межклеточные соединения, образованные выпячиваниями цитоплазмы одних клеток, вдающимися в цитоплазму других (см. рис. 1). За счет интердигитации увеличивается прочность соединения клеток друг с другом и нарастает площадь поверхности, через которую могут осуществляться межклеточные обменные процессы. (5) Щелевое соединение (nexus) образовано совокупностью трубчатых трансмембранных структур диаметром 9-11 нм (коннексонов), пронизывающих плазмолеммы соседних клеток на участках диаметром 0.5-3 мкм и стыкующихся друг с другом в области узкой межклеточной щели шириной 2-3 нм (см. рис. 1). Число коннексонов в щелевом соединении обычно исчисляется сотнями. Каждый коннексон представлен 6 (иногда 4 или 5) субъединицами, образованными белком коннексоном, и пронизан каналом диаметром 1.5-2.0 нм, который обусловливает свободный обмен низкомолекулярными (с массой до 2 кД) соединениями (неорганическими ионами, сахарами, витаминами, аминокислотами, нуклеотидами, АТФ и др.) между клетками, обеспечивая их ионное и метаболическое сопряжение. Базальная поверхность эпителиоцитов прилежит к базальной мембране, к которой она прикреплена с помощью полудесмосом - соединений, сходных по строению с половинами десмосом. Молекулярная организация и биохимический состав полудесмосом и десмосом, однако, не идентичны. В функциональном плане базальная и латеральная (до уровня плотных соединений) части плазмолеммы эпителиоцита в совокупности образуют единый комплекс (базолатеральную плазмолемму), который содержит специфические для него мембранные белки. Эти белки служат (а) рецепторами, воспринимающими различные сигнальные молекулы (гормоны, факторы роста), (б) переносчиками питательных веществ, поступающих из сосудов подлежащей соединительной ткани, (в) ионными насосами и др. Базальная поверхность может быть сравнительно плоской или образовывать выросты - базальные отростки. Последние могут служить для обеспечения более прочной связи эпителиоцитов с соединительной тканью путем увеличения площади поверхности их соприкосновения (например, в многослойных эпителиях).
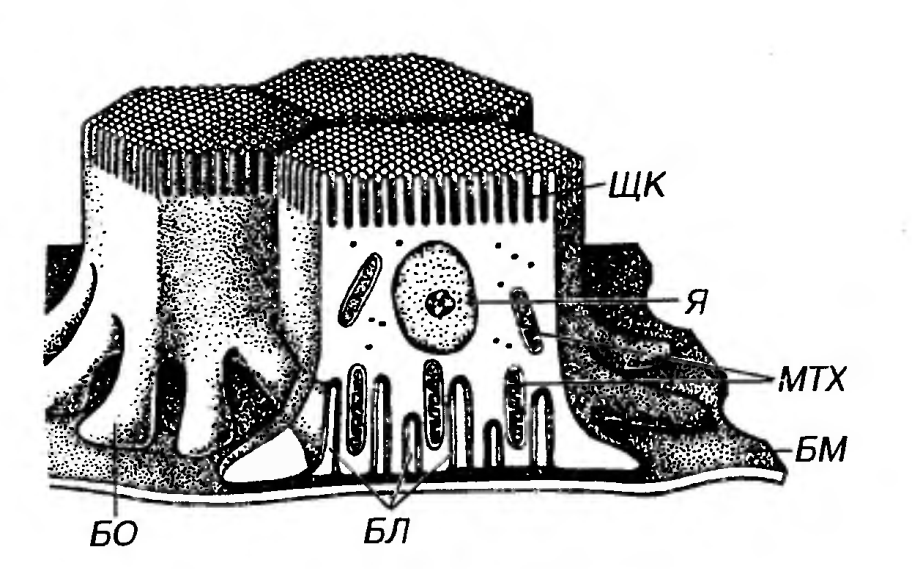
Базальная исчерченность или базальный лабиринт - термины, используемые для описания характерного строения базального отдела некоторых эпителиоцитов (например, в канальцах почки и части выводных протоков слюнных желез), образующих многочисленные базальные отростки, которые переплетаются друг с другом и с отростками других клеток (рис. 2). Цитоплазма таких базальных отростков содержит большое количество митохондрий, лежащих вдоль отростков (перпендикулярно базальной мембране) и вырабатывающих энергию, которая потребляется ионными насосами в их плазмолемме. Функция эпителиоцитов, обладающих базальной исчерченностью, связана с изменением ионного состава жидкости (мочи, слюны) в просвете указанных канальцев и протоков.
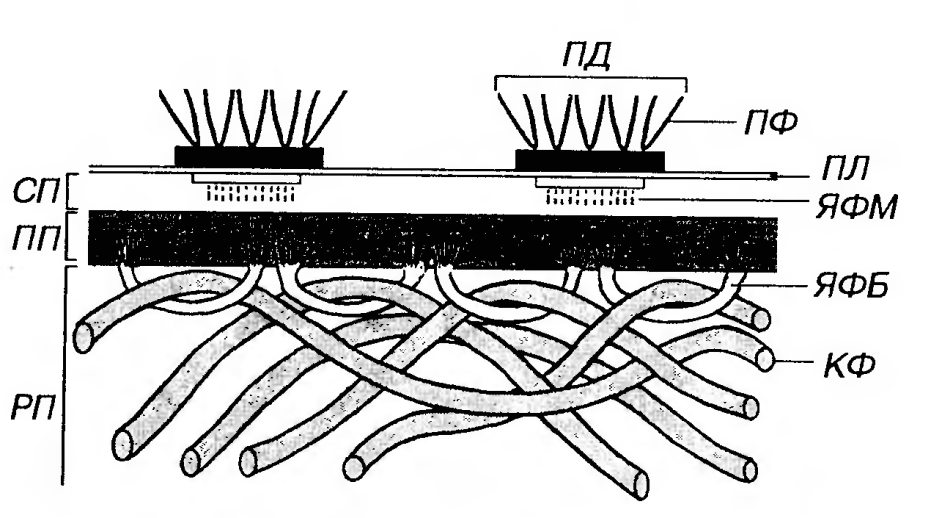
Рис. 2. Базальный лабиринт эпителиоцитов канальцев почки (проксимальный отдел нефрона). Я - ядро, ЩК - щеточная каемка, MTX - митохондрии, БМ - базальная мембрана, БЛ - базальный лабиринт, БО - базальные отростки. Базальная мембрана связывает эпителий и подлежащую соединительную ткань и образована компонентами, которые вырабатываются этими тканями. На светооптическом уровне на препаратах она имеет вид бесструктурной полоски, не окрашивается гематоксилином и эозином, выявляется солями серебра и дает интенсивную ШИК-реакпию. На ультраструктурном уровне (рис. 3) в базальной мембране описаны три слоя (в направлении от эпителия к соединительной ткани): (1) светлая пластинка, (2) плотная пластинка, (3) ретикулярная пластинка (последняя не всеми авторами рассматривается как компонент базальной мембраны).
Рис. 3. Ультраструктурная организация базальной мембраны эпителия. СП -светлая пластинка, ПП - плотная пластинка, РП - ретикулярная пластинка, ПЛ - плазмолемма, ПД - полудесмосома, ПФ - промежуточные филаменты, ЯФМ - якорные филаменты, ЯФБ - якорные фибриллы, КФ - коллагеновые фибриллы. Светлая пластинка (lamina lucida, или lamina r а r а) - светлый мелкозернистый слой толщиной 30-50 нм, прилежащий к плазмолемме базальной поверхности эпителиоцитов. От полудесмосом эпителиоцитов вглубь этой пластинки, пересекая ее, направляются тонкие якорные филаменты. Светлая пластинка содержит гликопротеины (в том числе сульфатированный гликопротеин ламинин) и антиген пузырчатки (способствующие прикреплению базальной части эпителиоцитов), а также протеогликаны (гепарансульфат). Плотная пластинка (lamina densa) - слой толщиной около 50-60 нм, образованный мелкозернистым или фибриллярным материалом, который располагается под светлой пластинкой и обращен в сторону соединительной ткани. В эту пластинку вплетаются якорные фибриллы, имеющие вид петель (образованы коллагеном VII типа), в которые продеты коллагеновые фибриллы подлежащей соединительной ткани. Плотная пластинка содержит коллаген IV типа, энтактин (сульфатированный гликопротеин, связывающий ламинин с коллагеном IV типа), гепарансульфат. В состав базальной мембраны входят также (непостоянно) коллаген V типа и адгезивный гликопротеин фибронектин. Ретикулярная (фиброретикулярная) пластинка (lamina reticu laris) состоит из коллагеновых фибрилл соединительной ткани, связанных с якорными фибриллами, и по толщине значительно превосходит светлую и плотную пластинки. В ее состав входят фибриллы, образованные коллагенами I и III типов (последний вид фибрилл именуют также ретикулярными). Хотя, по мнению некоторых авторов, эту пластинку не следует относить к собственно базальной мембране, именно она образует основную массу той структуры, которая выявляется ШИК-реакцией или окраской солями серебра и соответствует классическому описанию базальной мембраны на светооптическом уровне. Функции базальной мембраны: 1) поддержание нормальной архитектоники, дифференцировки и поляризации эпителия; 2) обеспечение прочной связи эпителия с подлежащей соединительной тканью. К базальной мембране прикрепляются, с одной стороны, эпителиальные клетки (с помощью полудесмосом), с другой - коллагеновые волокна соединительной ткани (посредством якорных фибрилл); 3) избирательная фильтрация питательных веществ, поступающих в эпителий (базальная мембрана играет роль молекулярного сита); 4) обеспечение и регуляция роста и движения эпителия по подлежащей соединительной ткани при его развитии или репаративной регенерации. В физиологических условиях базальная мембрана препятствует росту эпителия в сторону соединительной ткани. Это ингибирующее действие утрачивается при злокачественном росте, когда раковые клетки прорастают сквозь базальную мембрану в подлежащую соединительную ткань (инвазивный рост). Вместе с тем, прорастание базальной мембраны эпителиальными клетками выстилки сосудов (эндотелиоцитами) наблюдается и в норме при новообразовании сосудов (ангиогенезе). Нарушения строения и функции базальной мембраны часто обусловливают патологические изменения в органах. Ряд заболеваний почек - (гломерулонефритов) связан с повреждением базальной мембраны почечных клубочков (в которых происходит фильтрация крови с образованием мочи) вследствие иммунного повреждения, обусловленного антителами, клетками-эффекторами или активацией комплемента в ответ на отложение комплексов антиген-антитело. Утолщение базальной мембраны эндотелия мелких сосудов, отмечаемое при сахарном диабете, служит главной причиной их дисфункции (диабетической микроангиопатии). При этом резкое нарушение проницаемости сосудистой стенки вызывает дегенеративные процессы в различных органах (почках, сетчатке глаза, мышцах и др.). При пемфигоиде - одной из форм пузырчатки (см. выше) образуются аутоантитела к компонентам базальной мембраны, что вызывает разрушение последней и отделение эпителия от соединительной ткани с его гибелью и формированием подэпителиальных пузырей, давших название болезни. Апикальная поверхность эпителиоцитов может быть сравнительно гладкой или образовывать разнообразные вьшячивания. У некоторых эпителиоцитов на ней имеются специальные органеллы - микроворсинки и реснички. (1) микроворсинки - пальцевидные выросты цитоплазмы диаметром около 0.1 мкм и длиной до 1 мкм, основа которых образована пучком актиновых микрофиламентов, связанных как друг с другом, так и с внутренней поверхностью плазмолеммы (см. лекции по цитологии). Микроворсинки увеличивают площадь апикальной поверхности и в небольшом количестве могут встречаться на различных клетках. Они максимально развиты и многочисленны (до нескольких тысяч) в эпителиоцитах, участвующих в процессах всасывания (например, в тонкой кишке или канальцах проксимального отдела нефрона), где их совокупность называется щеточной (исчерченной) каемкой. Своеобразными вариантами микроворсинок являются так называемые стереоцилии и волоски. Стереоцилии (название отражает первоначальное ошибочное отнесение их к категории неподвижных ресничек) крупнее обычных микроворсинок (достигают в длину 5-7 мкм), могут ветвиться, истончаясь на концах, однако полностью соответствуют им по своей ультраструктурной организации. Они встречаются в эпителии некоторых участков семявыносящих путей (проток придатка яичка и семявыносящий проток). Предположительно участвуют в процессах всасывания жидкости, продуцируемой яичком. Волоски рецепторных сенсорно-эпителиальных (волосковых) клеток органов равновесия и слуха представляют собой видоизмененные микроворсинки (стереоцилии). Они не ветвятся, широко варьируют по длине (от 2 до 12 мкм в органе слуха и от 1 до 100 мкм в органе равновесия). Волоски имеют равномерную толщину (0.1-0.25 мкм) по всей длине, сужаясь у своего основания. Их цитоскелет образован актиновыми микрофиламентами, рыхло расположенными вокруг центрального плотного пучка филаментов, проникающего в виде корешка из волосков в апикальную цитоплазму клеток. Волоски участвуют в восприятии звука, гравитации и ускорений: их отклонение преобразуется в волну деполяризации рецепторных клеток. (2) реснички - крупные выпячивания цитоплазмы эпителиоцита диаметром порядка 0.2 мкм и длиной 5-10 мкм, основа которых образована аксонемой - каркасом из 10 пар микротрубочек, связанных с дополнительными белками (см. лекции по цитологии). Реснички являются органеллами движения; их синхронизированное биение осуществляется с частотой 10-25 колебаний/с в направлении, которое генетически предопределено природой эпителиоцита. Количество ресничек на апикальной поверхности одной клетки в различных эпителиях варьирует от нескольких десятков до нескольких сотен.Биение ресничек эпителия воздухоносных путей способствует перемещению по его поверхности и удалению слизи с прилипшими к ней микробами и частицами пыли. Повреждение или потеря ресничек вызывает нарушение этого важного защитного механизма очищения слизистой оболочки. Биение ресничек эпителия маточной трубы обусловливает транспорт ооцита сокружающими его фолликулярными клетками или эмбриона по направлению к матке.
|
||
|
|
Последнее изменение этой страницы: 2021-02-07; просмотров: 438; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.108 (0.012 с.) |