Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
II. Связь сновидений с глубиной снаСтр 1 из 12Следующая ⇒
Предисловие автора
В этой книге изложены наши представления о нервных механизмах сновидений. Предлагаемая физиологическая теория сновидений основана на учении акад. И. П. Павлова о высшей нервной деятельности и представляет собою попытку дать материалистическую теорию сновидений. Этот труд рассчитан на физиологов, психологов, врачей-клиницистов, в особенности психоневрологов, психиатров и невропатологов. Мы надеемся, что данная работа послужит началом для дальнейших исследований этого вопроса экспериментаторами и клиницистами.
Введение
Одним из источников всевозможных суеверий являются сновидения. Обывательские и суеверные представления о сновидениях, к сожалению, до сих пор широко распространены среди населения всего земного шара. Сновидения — психологический источник некоторых форм спиритуализма и мистицизма. Еще Ф. Энгельс указывал на сновидения как на один из источников первобытного анимизма. Прогресс научных знаний о сновидениях и распространение их постепенно вытеснят невежественные и мистические представления. Мы попытались изложить систему наших физиологических представлений о сновидениях на основе современной физиологии и патологии высшей нервной деятельности, созданной Иваном Петровичем Павловым и его школой. Нашей задачей является анализ нервных механизмов сновидений. Мы даем собственную концепцию на основе, главным образом, собственных фактических материалов и данных, полученных нашими сотрудниками. Поставленная задача имеет важное теоретическое и практическое значение. Мы полагаем, что наша цель будет достигнута, если нам удалось набросать хотя бы первоначальную схему физиологической теории сновидений. Существуют ли какие-либо научные предпосылки для физиологического понимания сновидений в трудах наших отечественных физиологов, занимавшихся исследованием деятельности головного мозга? Они существуют. Эти предпосылки мы находим в высказываниях И. М. Сеченова, И. П. Павлова, А. Ф. Самойлова, В. М. Бехтерева и др. Руководящей идеей нашей отечественной физиологии в этом отношении было — связать сновидения с деятельностью головного мозга во время сна и переходных состояний. Это положение является основным в нашей работе.
И в настоящее время ряд физиологов и клиницистов в СССР[1] и за границей пытается найти физиологические подходы к разрешению этого сложного и трудного вопроса. В СССР в течение последних лет вышло несколько научно-популярных работ (П. К. Анохин, Э. А. Асратян, С. И. Гальперин, А. С. Чистович, Л. Л. Васильев, Ф. П. Майоров, В. М. Касьянов, З. Косенко и др.), в которых также делались попытки дать физиологическое объяснение сновидениям. Наша лаборатория (Лаборатория физиологии и патологии высшей нервной деятельности человека Института физиологии им. И. П. Павлова) в течение последних 10 лет занималась систематическим исследованием проблемы сна и переходных состояний у людей в норме и патологии. Нами и сотрудниками был опубликован большой и новый научный материал, относящийся к динамике различных форм и фаз сна у человека. Наши экспериментальные исследования проводились в следующих трех направлениях: 1) исследование трех основных видов сна: естественного (ночного и дневного), гипнотического и наркотического; 2) сопоставление динамики естественного, нормального сна с динамикой различных форм патологического (нарколептического, истерического «летаргического» сна и др.); 3) сравнительно-физиологическое исследование онтогенетической эволюции процесса сна по возрастам (начиная от младенцев и кончая глубокими стариками). Вполне естественно, что при проведении такой работы мы должны были, следовательно, иметь дело со сновидениями и как с существенной стороной процесса сна, и как с неизбежным объектом наших исследований. Даже при стремлении оставаться на позиции объективного, физиологического изучения процесса сна как такового мы не могли бы, в целях более правильной оценки полученных результатов, игнорировать сновидения у человека. Может быть сон без сновидений, но не может быть сновидений без сна. Сновидения должно считать верным признаком сна. Но что же такое сновидения? Какова их сущность? В состоянии ли мы проникнуть в нервные механизмы сновидений и установить какие-либо закономерности? Таков стоящий перед нами комплекс вопросов. Предмет исследования весьма сложен. Сновидения — это наиболее запутанная и непонятная часть нашей психической, мозговой деятельности. Недаром некоторые ученые прошлого века приравнивали сновидения к «временному умопомешательству», а Каллен (Cullen) в XVIII в. считал сновидения «аналогичными бреду». Эти странные определения (как будет видно из дальнейшего изложения) имеют некоторый смысл. Действительно, в сновидениях мы наблюдаем самые беспардонные нарушения законов причинности, пространства, времени, морали и приличий, установленных воспитанием и социальными отношениями и т.п.
По поводу сновидений существует большая и разнообразная литература, написанная людьми самых различных специальностей. В этой литературе от древних веков до нашего времени имеется много ценного, но и не меньше заблуждений. Ответственной и специальной задачей будет разобраться в этом литературном материале и осветить с научной точки зрения историю вопроса. Но такая задача не составляет предмета нашей работы. Она сознательно оставляется нами в стороне. Мы даем изложение наших представлений без истории вопроса, без обзора и критики литературы. Это является минусом для монографического труда. Но мы и не претендуем на монографическое значение нашей книги. Это не будет означать того, что некоторые изложенные нами идеи и факты никогда не имели предшественников. Как уже было сказано, в содержании сновидений часто нарушаются представления о причинности, но сами сновидения подчинены этому основному закону, как все явления мира. Причинный анализ сновидений — одна из методологических предпосылок нашей работы. Другой методологической предпосылкой является эволюционно-физиологический подход к проблеме, т. е. сочетание эволюционного принципа с методом физиологического анализа на основе экспериментальных и клинико-физиологических данных павловской школы. Естественно, что мы исходили из признания реальности нашего субъективного мира, считая весьма важной задачей физиологической науки познание этого субъективного мира. Сновидения — это наиболее хаотическое проявление субъективного мира человека. В нашем изложении мы будем придерживаться принципа единства физиологии и психологии. В свое время И. П. Павлов формулировал как один из основных методологических принципов физиолога «принцип структурности», т. е. необходимость для физиолога постоянно связывать изучаемые им функции головного мозга с конкретными материальными структурами мозга. При физиологическом анализе сновидений это важное положение необходимо иметь в виду. Таковы основные методологические предпосылки нашей работы, исходящие из философии диалектического материализма. При исследовании высшей нервной деятельности человека кроме регистрации условных и безусловных рефлексов существенно важно также учитывать словесный отчет испытуемых относительно афферентной и эфферентной сторон этих рефлексов. Проводимые таким способом в нашей и других лабораториях опыты показали важное значение «синтеза субъективного и объективного» (по И. П. Павлову) для более глубокого понимания закономерностей высшей нервной деятельности человека. И. П. Павлов стремился понять собственную психическую деятельность на основе данных созданной им физиологии высшей нервной деятельности. Он говорил: «По крайней мере я, глядя на эти опыты, многое уяснил себе в себе и в других» (Полн. собр. трудов, т. III, 1949, стр. 390).
И. П. Павлов имел намерение дать физиологическое описание и физиологический анализ собственных сновидений. Наш фактический материал состоит: 1) из данных лабораторных экспериментов по изучению сна и сновидений, 2) из клинических наблюдений и 3) из данных жизненных наблюдений и собственных сновидений. Весь этот материал был собран нами в течение нескольких лет. Излагаемая здесь система представлений о физиологической сущности и нервных механизмах сновидений есть плод многолетних наблюдений и размышлений физиолога, посвятившего себя исследованию деятельности головного мозга. При изложении наших взглядов нам придется обращаться к психологическому описанию фактов и их физиологическому анализу на основе экспериментальных и клинических данных павловской школы. Мы будем стремиться к тому, чтобы дать не только физиологическое объяснение фактам, но и, по возможности, сделать физиологические выводы из них.
I. Нервный механизм сна
Наши представления о нервных механизмах сновидений опираются на теорию сна И. П. Павлова и на экспериментальные данные его школы. Поэтому мы начинаем изложение с представлений И. П. Павлова о физиологической сущности сна. На основании многочисленных опытов с условными рефлексами И. П. Павлов пришел к выводу, что в основе процесса сна лежит нервный механизм иррадиации торможения в коре больших полушарий с последующим распространением торможения на подкорковые области головного мозга. Этот нервный механизм является ведущим звеном в сложном физиологическом процессе сна. Исследования П. К. Анохина (1939) на двух сросшихся человеческих близнецах доказали преобладание нервного механизма сна над гуморальным. Близнецы имели общее туловище и две головы, одно сердце и общую кровеносную систему. Оказалось, что когда одна голова спала, то другая могла бодрствовать. Это значит, что один головной мозг мог находиться в состоянии сна, а другой — нет, несмотря на общие для них гуморальные условия. Это не значит, что гуморальный механизм не играет роли в развитии сна. Он имеет существенное значение, хотя у высших животных и человека отходит на второй план. Иррадиированное торможение, лежащее в основе сна, Павлов назвал «сонным торможением». Опытами на собаках сначала было установлено, что внутреннее корковое торможение и сон есть один и тот же процесс. Если происходит иррадиация какого-либо внутреннего торможения (угасательного, дифференцировочного, запаздывательного), то развивается сон. Приведем пример с угашением. Обычно в павловских лабораториях условные раздражители сопровождаются пищевым подкреплением. Если какой-либо выработанный условный пищевой раздражитель (например звонок) повторять в опыте через одинаковые интервалы и не сопровождать едой, то условный слюнной рефлекс постепенно уменьшается и доходит до нуля, как мы говорим, «угасает». В основе угасания лежит активный тормозный процесс, задерживающий работу соответствующих корковых и подкорковых центров и связанных с ними периферических аппаратов (в данном случае слюнной железы). Если продолжать далее угашать условный рефлекс на звонок, то животное может заснуть (закроет глаза, опустит голову, повиснет в лямках, начнет храпеть). Это объясняется распространением, иррадиацией угасательного торможения из коркового слухового центра сначала по коре, а потом в подкорку. Закономерность иррадиации тормозного процесса в пространстве и во времени (а также и возбуждения) была доказана многочисленными экспериментами павловской школы. Процесс иррадиации торможения происходит волнообразно, с колебаниями.
В опытах на кожном анализаторе можно было проследить иррадиацию торможения из одного тормозного пункта — очага — в другие положительные, находящиеся на различном расстоянии от первого. В этих опытах также можно было проследить и обратный процесс — процесс концентрации, сосредоточения торможения в тормозном пункте. Следствием концентрации торможения в каком-то корковом очаге может быть положительная индукция по периферии данного очага. Положительная индукция усиливает процесс коркового возбуждения и устраняет сон. Почему же в корковых нервных клетках появляется торможение? Это происходит вследствие основного закона их деятельности — закона взаимодействия возбуждения и торможения. При известных условиях возбуждение сменяется торможением, а торможение, в свою очередь, — возбуждением. Если действует условный положительный раздражитель и не подкрепляется, то развивается торможение. Если действует подкрепляемый, но монотонный, слабый или чрезмерно сильный, условный раздражитель, то также возникает торможение. Это торможение является коренным функциональным свойством корковых нервных клеток и имеет, по Павлову, «охранительное» значение: оно охраняет клетки от возможного функционального истощения и разрушения; кроме того, во время развития торможения происходит восстановление работоспособности корковых клеток. «Сонное торможение» также имеет функцию «охранительного торможения» и способствует восстановлению работоспособности коры больших полушарий. Чем глубже сон, т. е. чем интенсивнее и экстенсивнее сонное торможение, тем больше л полнее восстановление работоспособности.
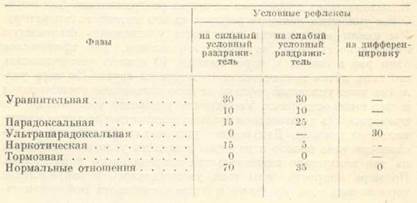
Сонное торможение как иррадиированная форма коркового торможения может возникать из любой формы торможения, появляющегося в коре больших полушарий. Сонное торможение может развиваться как из форм коркового выработанного, внутреннего, «условного торможения» (угасательного, дифференцировочного, запаздывательного и условного тормоза), так и из форм коркового врожденного, «безусловного торможения» (внешнего торможения, т. е. отрицательной индукции, и запредельного торможения), а также из комбинации того и другого. Эта физиологическая схема действительна как в отношении животных, так и в отношении людей. Сон при отрицательной индукции у собак был описан в работе К. С. Абуладзе (1949). При повышении пищевой или половой возбудимости у собак, а также при действии сверхсильных условных раздражителей может развиваться запредельное торможение, вызывающее уменьшение условных рефлексов и гипнотические фазы, которые И. П. Павлов рассматривал как «переходные состояния» между бодрствованием и сном. У животных и людей запредельное торможение является одним из нервных механизмов гипноза и гипнотических состояний. Запредельное торможение так названо Павловым потому, что оно вмешивается в деятельность коры каждый раз, когда корковое возбуждение (под влиянием внутренних или внешних условий) доходит до своего функционального предела. В этом случае прогрессивное увеличение условных рефлексов сменяется их уменьшением, иногда прогрессивным падением до нуля, т. е. до полного тормозного эффекта. Для иллюстрации этого важного положения приведем два примера из работ лаборатории И. П. Павлова. В. В. Рикман в 1930–1931 гг. проводил опыты с хроническим неполным голоданием на нескольких собаках разного типа нервной системы (Ф. П. Майоров, 1948а, стр. 252). По мере голодания у собак увеличивалась пищевая возбудимость, росли условные рефлексы; слабые условные раздражители стали давать эффект, равный сильным, дифференцировки растормозились. Увеличивавшиеся изо дня в день условные рефлексы достигли известного предела, за которым дальнейшее голодание повело уже не к увеличению, а к катастрофическому падению условных рефлексов. Это падение прогрессировало с продолжением голодания. Нами были поставлены следующие опыты (Майоров, 1940). В системе разных условных пищевых раздражителей был выработан условный пищевой рефлекс на тон катодного генератора. Мы имели возможность постепенно увеличивать физическую интенсивность тона, не меняя его высоты. Тон применялся один раз в опыте, и через каждые 10 опытов увеличивалась его сила путем незначительного прироста звука. По мере усиления тона условный рефлекс на него постепенно увеличивался и стал выше остальных. Увеличение условного рефлекса на тон продолжалось до известной его интенсивности и достигло максимального предела. Дальнейшее увеличение физической интенсивности тона повело к падению величины условного рефлекса на тон вследствие вмешательства запредельного торможения. По мере прироста силы тона увеличивалось и запредельное торможение, т. е. все более и более уменьшался рефлекс и дошел до нуля. Предел работоспособности коры больших полушарий различен у разных типов нервной системы: у сильных типов он выше, чем у слабых. При истощении сильной нервной системы понижается предел ее работоспособности. У животных и людей слабого типа наблюдается более частое вмешательство запредельного торможения и развитие гипнотических фаз и сна на основе механизма запредельного торможения. В основе частых и внезапных припадков сна при нарколепсии, надо думать, лежит хроническое истощение коры больших полушарий, резкое снижение предела ее работоспособности и вследствие этого — частое вмешательство запредельного торможения, вызывающего нарколептический сон. Переход от бодрственного состояния ко сну и от сна к бодрственному состоянию совершается через ряд гипнотических фаз. Эти фазы могут встречаться и в процессе самого сна, когда он неглубок. Гипнотические фазы могут возникать также при разных патологических состояниях больших полушарий головного мозга. Мы их опишем здесь, так как они нам пригодятся в дальнейшем. Нормальные отношения в опытах с условными рефлексами обусловливаются законом зависимости величины условного рефлекса от силы условного раздражителя. Сильным условным раздражителям соответствуют большие условные рефлексы, слабым условным раздражителям соответствуют меньшие условные рефлексы, тормозные раздражители (дифференцировка, условный тормоз) вызывают полный (нулевой) или неполный тормозный эффект. Эти нормальные отношения нарушаются при развитии гипнотических фаз. Основные гипнотические фазы следующие:
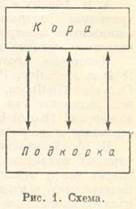
В этой таблице приведены схематические, примерные, отношения между условными рефлексами при гипнотических фазах. Для сравнения даны нормальные отношения. Уравнительная фаза характеризуется уравнительными отношениями между условными рефлексами на сильные и слабые раздражители: рефлексы выравниваются на разных уровнях. При парадоксальной фазе условные рефлексы на сильные раздражители делаются меньше рефлексов на слабые раздражители; слабые же раздражители могут давать эффект и больший, чем в норме. При ультрапарадоксальной фазе извращаются отношения между положительным и тормозным (дифференцировочным) раздражителями, например: условный рефлекс на положительный раздражитель (стук метронома частотой в 120 ударов за 1 мин.) вместо обычного эффекта в 70 делений шкалы дает нуль (тормозный эффект), а дифференцировочный раздражитель (стук метронома частотой в 60 ударов за 1 мин.) вместо обычного нулевого эффекта вызывает положительный эффект 30. Наркотическая фаза характеризуется снижением условных рефлексов на сильные и слабые раздражители при более значительном падении эффекта слабых раздражителей. Тормозная фаза есть фаза уже полного торможения. Между описанными основными гипнотическими фазами могут встречаться промежуточные. На них нам нет надобности останавливаться. Учение И. П. Павлова о гипнотических фазах представляет важное физиологическое основание для понимания нервных механизмов сновидений. Переходим к существенному для нас вопросу о растормаживании. Всякий корковый тормозный эффект при известных условиях может быть расторможен. Так, дифференцировка, обычно дающая нулевой эффект, может быть расторможена при повышении корковой возбудимости или при действии постороннего раздражителя, или при каком-либо другом обстоятельстве. При развитии сна, когда сонным торможением захватываются большие районы коры и подкорки, могут происходить растормаживания отдельных очагов под влиянием фазовых колебаний возбудимости в коре или под влиянием афферентных импульсов, поступающих в кору от внешних и внутренних рецепторов. Тогда на фоне разлитого торможения в коре могут возникать, исчезать и снова возникать отдельные, хаотически разбросанные очаги возбуждения. Механизм растормаживания коры и подкорки в процессе сна имеет важное значение для физиологического понимания сновидений. Корковая деятельность постоянно связана с колебаниями возбудимости, тонуса коры больших полушарий. Понижение возбудимости коры способствует развитию сонного торможения. Нормально высокий тонус коры во многих отношениях зависит от постоянного притока афферентных импульсов от внешних и внутренних рецепторов — от системы экстероцепторов, с одной стороны, и системы интероцепторов и проприоцепторов, с другой. Известно наблюдение Штрюмпеля: у одного больного из дистантных органов чувств остались действующими только один глаз и одно ухо. Когда этому больному закрывали действующие глаз и ухо, то он погружался в сон. Еще ранее Штрюмпеля аналогичный случай наблюдал и описал С. П. Боткин. Его больная была лишена всех видов чувствительности за исключением кожной и мышечной. Обычно больная была погружена в сон и пробуждалась лишь при прикосновении к руке, сохранившей чувствительность. В этом отношении важное значение имеют опыты на собаках, лишенных трех дистантных рецепторов (зрительного, слухового и обонятельного), начатые В. С. Галкиным в лаборатории А. Д. Сперанского. В. С. Галкин получил такую собаку, которая находилась в состоянии почти постоянной спячки. Она просыпалась лишь под влиянием голода и раздражений с прямой кишки и мочевого пузыря. К. С. Абуладзе (1935) продолжил и углубил эти исследования. Его собака отличалась резким понижением корковой возбудимости, реактивности и чрезвычайной наклонностью ко сну. Из двух практиковавшихся условных рефлексов, пищевого и кислотного, мозговая кора такого животного могла осуществлять только один, а другой рефлекс, по закону отрицательной индукции, подвергался торможению. И. П. Павлов по этому поводу говорил, что при таком снижении реактивности коры у последней как бы не хватало энергии на два рефлекса. Тонус коры больших полушарий постоянно колеблется, находясь в зависимости от противоположных влияний — возбуждающих и гипнозогенных (снотворных). При развитии сна необходимо учесть происходящее при этом изменение во взаимоотношении системы экстероцепторов и системы интеро-проприоцепторов. Первая может быть заторможена и выключена, в то время как вторая будет продолжать действовать и посылать потоки афферентных импульсов от внутренних органов в кору. При дальнейшем углублении сна и висцеральные афферентные импульсы оказываются ниже порога возбудимости коры. Возбудимость коры также зависит от возбудимости подкорки. Подкорковое возбуждение сообщается коре, и кора в свою очередь может тонизировать подкорку. Высшая нервная деятельность представляет взаимодействие коры и подкорки. Вне этого основного взаимодействия не протекает ни одно явление высшей нервной деятельности, в том числе и сон во всех его видах и стадиях. Взаимодействие коры и подкорки (рис. 1) может быть как возбуждающего, так: и тормозящего характера. Важное значение, кроме прямой импульсации, здесь имеет взаимная индукция: возбужденное состояние коры при известных условиях может вызвать отрицательную индукцию в подкорке (т. е. произвести в ней торможение); тормозное состояние коры может вызвать положительную индукцию в подкорке (т. е. повысить в ней возбуждение); иногда сильное возбуждение подкорковой эмоциональной, аффективной деятельности вызывает торможение коры по закону отрицательной индукции. При развитии сонного торможения только в коре больших полушарий наблюдается стадия подкорковой положительной индукции. При иррадиации сонного торможения с коры на подкорку затухает подкорковое возбуждение, наступает более глубокая стадия сна.
Предложенная И. П. Павловым теория сна может быть названа корковой теорией сна. Торможение возникает в коре, иррадиирует сначала по коре, а потом захватывает подкорковые области. Процесс распространения сонного торможения проходит ряд многообразных фаз и состояний, о которых мы уже говорили и которые нам придется рассмотреть в дальнейшем. Корковое происхождение сна, по Павлову, надо понимать на основе взаимодействия коры и подкорки. Среди некоторых клиницистов и физиологов распространено представление о «подкорковом центре сна» [Экономо (Economo, 1929); Гесс (Hess, 1931); Роже (Roger, 1946); Лермитт (Lhermitte, 1948); Рубино (Rubino, 1949), и др.]. Н. Клейтман (N. Kleitman, 1939) считает, что в подкорковой области существует специальный «центр бодрствования», от деятельности которого зависит смена бодрствования и сна. Лермитт и Турнэй (Lhermitte et Tournay, 1927) признают существование двух противоположных центров на дне третьего желудочка — центра сна и центра бодрствования («dispositif, régulateur du sommeil et de la veille»). Ряд клинических наблюдений развития сонливости и сна у больных эпидемическим (летаргическим) энцефалитом и при опухолях стволовой части головного мозга привел некоторых невропатологов (Economo, 1929, и др.) к заключению о существовании «подкоркового центра сна». Гесс (Hess, 1931) вызывал сон у подопытных кошек с вживленными электродами в задней части таламической области электрическим раздражением. На основании таких экспериментов Гесс заключил, что в этом месте мозга локализируется «подкорковый центр сна». Гесс (Hess, 1933) также вызывал сон у животных введением в область третьего желудочка эрготамина. Демоль (Demole, 1927) вводил кальций в область tuber cinereum и получал сон. А. В. Тонких и Е. А. Моисеев получили сон у кошек электрическим раздражением гипоталамической области через вживленные электроды. Но они не считают этот факт доказательством наличия центра сна в указанной области. Некоторые исследователи [например, Фултон и Бейли (Fulton a. Bailey, 1929); Наута (Nauta, 1946); Бронк (Вrоnс); Гельхорн (Gellhorn, 1943)] считают роль гипоталамуса в нервном механизме сна весьма существенной. В последнее время Конорский (Konorski, 1948) также считает возможным существование подкоркового центра сна. Н. А. Рожанский (1949) полагает, что существуют два нервных механизма сна — корковый и подкорковый. Разногласия между отдельными авторами в отношении локализации подкоркового центра сна он объясняет тем, что в подкорковой области имеется множественная система центров сна и центров бодрствования. Рожанский придает большое значение рефлекторным механизмам, действующим с сетчатки глаза: так, у дневных птиц колбочковый аппарат обусловливает бодрственное состояние, а палочковый — сонное, у ночных птиц — наоборот. Чем выше в эволюционном ряду стоит какой-либо вид животных, тем большее значение приобретает у него корковый механизм сна, особенно у человека. Таково «согласование» корковых и подкорковых механизмов сна, предложенное Н. А. Рожанским. Мы полагаем, что колбочковый и палочковый механизм сна—бодрствования не может иметь универсального значения. Что же касается концепций Конорского и Рожанского, то надо признать, что они являются ошибочными и опровергаются павловской теорией сна. Таким образом, имеются многочисленные клинические и экспериментальные данные, на основании чего некоторые пришли к заключению о существовании специального подкоркового центра сна в области дна третьего желудочка мозга и задней части таламо-гипоталамической области. Однако И. П. Павлов в докладе в 1932 г. дал вполне убедительное физиологическое объяснение приведенным данным, исходя из своей корковой теории сна. Факты, якобы говорящие за теорию подкоркового центра сна, он объяснил как случаи патологического и экспериментального (посредством блокады) выключения интероцептивных и проприоцептивных импульсов, обычно поддерживающих соответствующий корковый тонус. Тогда происходит падение корковой возбудимости, что благоприятствует развитию сонного торможения в коре и дальнейшей его иррадиации на подкорку. Следовательно, все имеющиеся клинические и экспериментальные факты сна могут быть поняты с единой физиологической точки зрения, предложенной Павловым. С этой точки зрения распространение процесса сна происходит с коры больших полушарий на подкорку, а не наоборот, с диэнцефалона на кору, как полагает, например, Анри Роже (Н. Roger, 1946). Неоспоримое преимущество павловской теории сна заключается в том, что она объясняет как факты коркового происхождения сна (многочисленные данные павловской школы), так и факты так называемого «подкоркового происхождения» сна. Теорией же подкоркового центра сна пытаются объяснить лишь одну группу фактов «подкоркового происхождения» сна, и она беспомощна в объяснении фактов коркового происхождения сна. Как можно было бы объяснить при помощи теории подкоркового центра сна хотя бы один элементарный и весьма распространенный в опытах лабораторий Павлова факт развития сна у собак вследствие угашения условного рефлекса? Однако имеются экспериментальные факты, которые как будто бы действительно находятся в противоречии с корковой теорией сна. У собак и кошек без больших полушарий обнаруживается чередование сна и бодрствования. При этом наблюдается преобладание периода сна над периодом бодрствования. И. П. Павлов предполагал существование двух форм сна: сна «активного» (в случае развития разлитого торможения в коре и подкорке) и сна «пассивного» (в случае резкого падения возбудимости при выключении дистантных рецепторов или при удалении коры больших полушарий). По этому поводу необходимо высказать следующее соображение. Согласно представлениям И. П. Павлова, как возбуждение, так и торможение суть функции каждого нервного центра и каждой нервной клетки. Торможение может возникать в любом комплексе нервных клеток и иррадиировать на другие; это может иметь место и в подкорке у бесполушарных животных, поэтому и в последнем случае нет надобности признавать какой-либо специальный подкорковый центр сна. Этот вопрос требует дальнейших экспериментальных исследований. Переходим к краткому рассмотрению вопроса о роли вегетативной нервной системы в развитии сна. Раньше полагали, что сон есть «царство вагуса». Это положение теперь уже должно быть лишено своего категорического характера. Симпатическая нервная система также играет существенную роль в развитии сна. Так, опытами А. В. Тонких (1940) было доказано, что электрическое раздражение гипоталамической области вызывает у кошек сон в том случае, если сохранена симпатическая иннервация головы. В случае оперативного нарушения симпатических связей электрическое раздражение указанной области не вызывало сна у кошек. Мы, совместно с М. И. Неменовым и Л. С. Васильевой (1949), производили облучение рентгеновскими лучами шейных симпатических узлов у собак, в результате чего у животных наблюдалось развитие сонливости и сна в экспериментальной камере для условных рефлексов. Здесь можно предполагать как прямое действие с шейных симпатических узлов на головной мозг и его сосуды, так и косвенное — через переднюю долю гипофиза. Имеются клинические данные об участии гипофиза в явлениях сна (Сальмон и др.). Эксперименты А. В. Тонких также показали, что электрическое раздражение гипоталамической области через вживленные электроды не вызывает сна у кошек, если у них удален гипофиз. В физиологии установлен также следующий факт. Если у животных вызвать сон при помощи каких-либо снотворных (хлоралгидрата, уретана и др.), то такой сон может быть прекращен введением адреналина и других симпатомиметических веществ. В связи с этим необходимо напомнить данные А. В. Тонких (1938): парасимпатические вещества (ацетилхолин и пилокарпин) удлиняли «гипноз» (каталептоидное состояние) у лягушек, адреналин же прекращал это состояние. Таким образом, известное взаимодействие парасимпатической и симпатической нервной системы и эндокринных органов (в частности гипофиза) имеет существенное значение в физиологическом механизме сна. При развитии сна происходит функциональная перестройка в процессах вегетативной нервной системы и системы эндокринной. Эта перестройка тесно связана с ведущим нервным механизмом сна — иррадиацией сонного торможения в коре и подкорке. В свою очередь наклонность к развитию диффузного торможения в коре больших полушарий зависит от вегетативных влияний на кору. Изложенными представлениями о нервном механизме сна не исчерпывается вопрос о физиологической природе сна. Основой самого нервного или нервно-гуморального механизма сна является химизм. В свое время в физиологической науке была распространена токсическая теория сна в различных ее вариантах [Лежандр (Legendre, 1911); Пиерон (Piéron, 1913); Клапаред (Claparède, 1905, 1928); Демоль (Demole, 1927), и др.]. Предполагалось, что в результате физической и умственной работы в организме появляются гипотетические яды (кенотоксин, гипнотоксин и т.п.), которые и вызывают развитие сна. Разные авторы давали разные названия этим «снотворным» веществам — продуктам утомления. Биологические эксперименты, проведенные этими авторами, указывали на химические изменения, происходившие в организме животных при развитии сна. Но истинная химическая основа сна оставалась и остается до сих пор невыясненной. Тут играет известную роль баланс в содержании калия и кальция в крови и цереброспинальной жидкости, содержание в крови брома и других веществ. И. П. Павлов и его сотрудники много занимались исследованием действия солей брома на высшую нервную деятельность. Этими опытами было установлено, что бром способствует усилению процесса торможения. Опытами М. Ф. Васильева (1948) было доказано уменьшение содержания брома в крови у собак при старении. Цондек и Бир (Zondek u. Bier, 1932) на основании своих исследований считали, что у старых людей уменьшается содержание брома в гипофизе. Исходя из приведенных данных о броме, мы сделали предположение, что ослабление функции сна у стариков надо поставить в связь с возможным уменьшением у них брома в крови и в тканях. Нашему сотруднику А. Л. Саперу в 1940 г. было поручено провести опыт лечения старческой бессонницы хроническим бромированием малыми дозами. Работа была проведена в Ленинграде в одном из домов для престарелых инвалидов на 50 стариках (в возрасте от 60 до 95 лет). Под наблюдением были лица со старческим ослаблением сна (в смысле его глубины и продолжительности), но без какой-либо серьезной органической основы. Бром применялся в виде 1%-го раствора бромистого натрия ежедневно в течение нескольких недель и месяцев по 1 столовой ложке 3 раза в день. Таким образом, за сутки эта доза составляла 0.45 Na-Br. У большинства был получен положительный эффект — углубление и удлинение сна. Те лица, которые не засыпали днем после обеда, стали в это время спать. Положительный эффект обычно наступал на 7–10-й день от начала бромирования. Таким образом, при помощи брома можно было усилить сонное торможение. Однако вопрос о химической основе сна остается вопросом будущего. Нам остается еще сказать несколько слов о наркотическом сне и рассмотреть вопрос о сущности гипноза.
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-02-07; просмотров: 118; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.133.161.153 (0.056 с.) |