Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
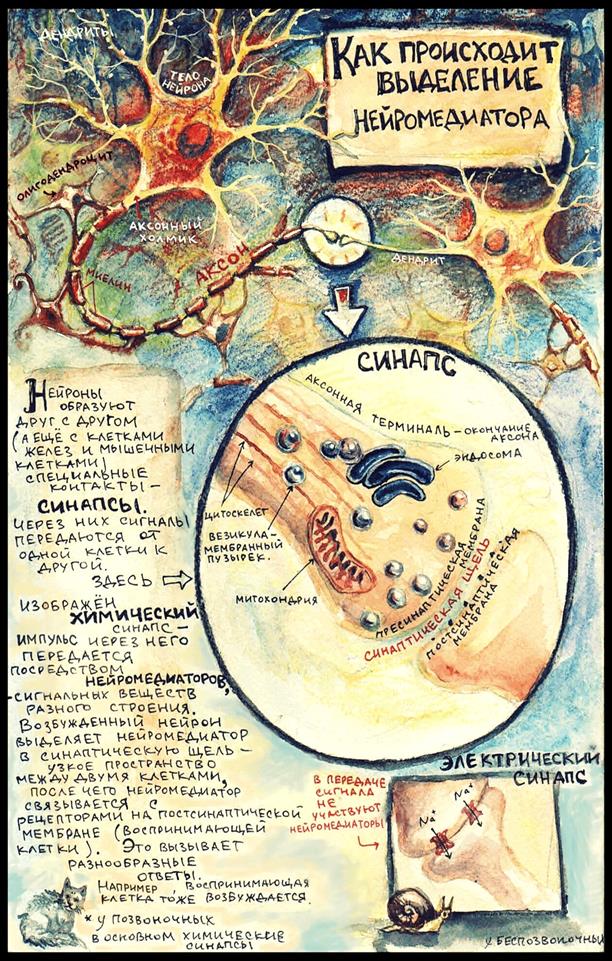
Способы регуляции синаптической передачи (синаптическое облегчение и синаптическая депрессия)
1.Синаптическая депрессия - это ухудшение проведения в синапсах в результате длительной посылки импульсов, например, при длительном раздражении афферентного нерва (утомляемость центра). Утомляемость нервных центров продемонстрировал Н. Е. Введенский в опыте на препарате лягушки при многократном рефлекторном вызове сокращения икроножной мышцы. В этом случае ритмическое раздражение одного нерва вызывает ритмические сокращения мышцы, приводящие к ослаблению силы ее сокращения вплоть до полного отсутствия сокращения. Переключение раздражения на другой нерв сразу же вызывает сокращение той же мышцы, что свидетельствует о локализации утомления не в мышце, а в центральной части рефлекторной дуги. 2.Синаптическое облегчение - это улучшение проведения в синапсах после короткого раздражения афферентных путей. Степень выраженности облегчения возрастает с увеличением частоты импульсов, оно максимально, когда импульсы поступают с интервалом в несколько миллисекунд.
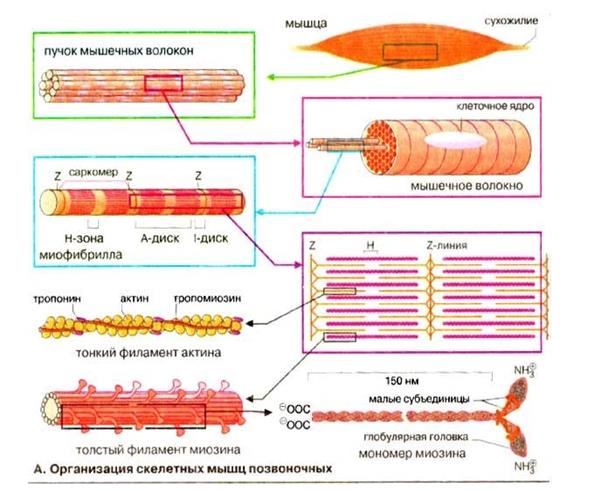
Скелетная мышца: функциональное значение отдельных структурных элементов мышечного волокна, понятие о структурной и функциональной единице изолированной мышцы и двигательного аппарата организма, классификация двигательных единиц Скелетная мышца состоит из множества мышечных волокон, имеющих точки прикрепления к костям и расположенных параллельно друг другу. Каждое мышечное волокно (миоцит) включает множество субъединиц – миофибрилл, которые построены из повторяющихся в продольном направлении блоков (саркомеров). Саркомер является функциональной единицей сократительного аппарата скелетной мышцы. Миофибриллы в мышечном волокне лежат таким образом, что расположение саркомеров в них совпадает. Это создает картину поперечной исчерченности.
Толстые филаменты состоят из белка миозина, тонкие-из актина, тропопнина и тропомиозина. Миозиновые филламенты образованы повторяющимися молекулами белка миозина. Каждая молекула миозина имеет головку и хвост. Головка миозина может связываться с молекулой актина, образуя так называемый поперечный мостик.
Клеточная мембрана мышечного волокна образует инвагинации (поперечные трубочки), которые выполняют функцию проведения возбуждения к мембране саркоплазматического ретикулума. Саркоплазматичекий ретикулум (продольные трубочки) представляет собой внутриклеточную сеть замкнутых трубочек и выполняет функцию депонирования ионов Са++
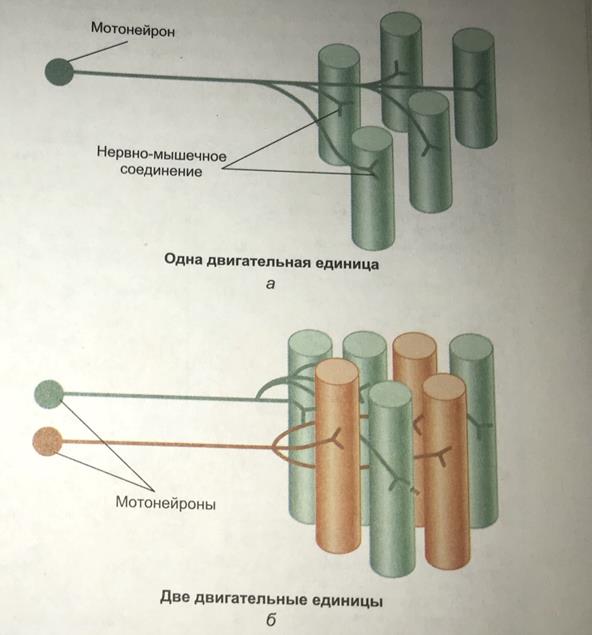
Светлая(изотропная)полоса-I-диск, расположена между краями А-дисков двух соседних саркомеров и состоит из тех участков, которые не перекрываются толстыми филаментами. Z-пластинка делит I-диск пополам. Волокна скелетных мышц иннервируются аксонами нервных клеток--мотонейронами(или соматическими эфферентными нейронами). Тела этих клеток расположены в спинном мозге или в стволе мозга. Мотонейрон и мышечные волокна, которые он иннервирует, составляют двигательную единицу.
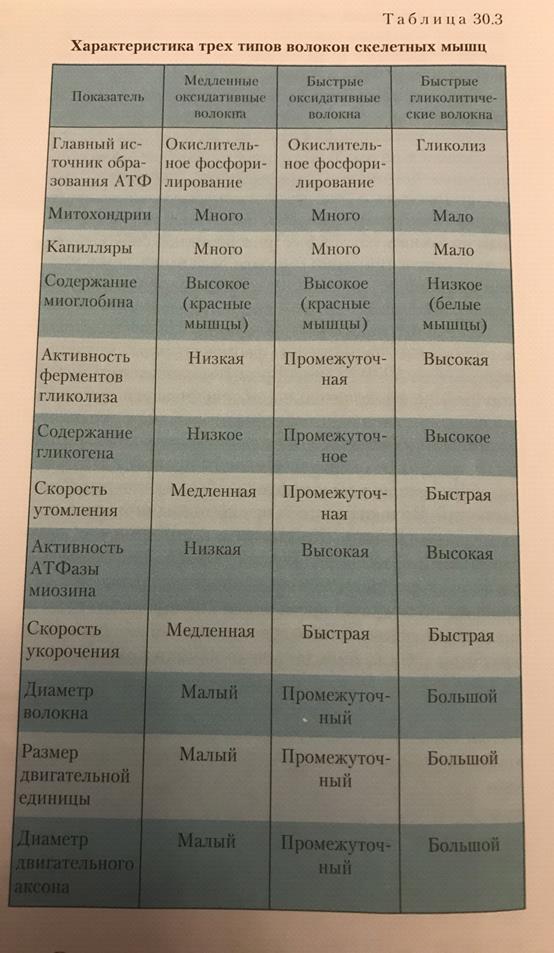
Область плазматической мембраны, лежащая непосредственно под нервным окончанием, обладает особыми свойствами и называется двигательной концевой пластинкой. Структура, состоящая из нервного окнчания и двигательной концевой пластинки, --это нервно-мышечное окончание(нервно-мышечный синапс). Типы двигательных единиц: -Имеют относительно большой диаметр -Отличаются светлым цветом, поскольку в них снижено количество миоглобина -Окружены небольшим количеством капилляров -Имеют относительно небольшое количество митохондрий -Характеризуются высоким содержанием гликогена
Получают АТФ за счет гликолиза. Из-за небольшого количества капилляров и миоглобина получают мало кислорода. Незначительно количество митохондрий в цитозоле также определяет небольшое потребление кислорода. Высокое содержание гликогена обуславливает достаточное количество глюкозы для гликолиза. Посему такие волокна называют гликолитическими. Красные мышечные волокна: -Диаметр красных мышечных волокон примерно в 2 раза меньше диаметра белых -Темно-красный цвет из-за высокого содержания миоглобина -Окружены большим количеством капилляров -Имеют множество митохондрий -Низкое содержание гликогена Для синтеза АТФ используют цикл Кребса и окислительное фосфорилирование. Небольшой диаметр капилляров облегчает диффузию кислорода. Из-за небольшого содержания гликогена для энергетического обеспечения красных волокон используются жирные кислоты. Они распадаются до ацетил КоА и включатся в цикл Кребса. Посему они называются оксидативными.
1. Медленные оксидативные(тип1)-низкая активность миозиновой АТФазы и высокая окислительная способность. 2.Быстрые оксидативные(тип2а)-высокая активность миозиновой АТФазы и высокая гликолитическая способность. 3.Быстрые гликолитические волокна(тип2б)-высокая активность миозиновой АТФазы и высокая гликолитическая способность.
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-01-08; просмотров: 1758; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.226.222.12 (0.006 с.) |