Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Регуляция по времени и развитие крыла цыпленкаСодержание книги
Поиск на нашем сайте
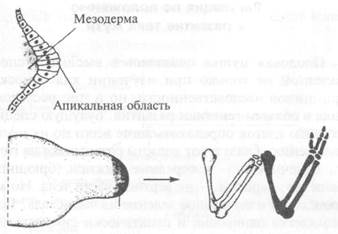
Прекрасный пример временного механизма — развитие крыла цыпленка (рис. 11.2). Крыло вырастает из задатка конечности, состоящего из клеток мезодермы, покрытых слоем эктодермы, включая апикальную область — растущий кончик задатка. Клетки этого кончика посылают инструкции клеткам мезодермы, и, вероятно, в них имеются какие-то внутренние часы, определяющие порядок инструкций. Дж. X. Льюис, Д. Саммербелл и Льюис Уол-перт проводили эксперименты по удалению кончика и перемещению его на задатки другого возраста. Если переместить кончик, находящийся на более ранней стадии развития, в задаток, в котором уже сформировались плечевая, лучевая и локтевая кости, то кончик посылает инструкции по повторному формированию этих костей. Но если переместить кончик более поздней стадии на задаток ранней стадии, то он будет посылать инструкции по формированию конечных фаланг. Этот эксперимент показывает, что часы апикальной области сначала включают гены, определяющие строение плечевой кости, затем эти гены выключаются и включаются гены формирования лучевой и локтевой костей. После этого они тоже выключаются и включаются гены фаланг. Рис. 11.2. Нормальное крыло птенца содержит набор костей, обычный для всех позвоночных: плечевая кость (верхняя часть конечности), лучевая и локтевая кости (нижняя часть), кости запястья и фаланги (пальцы). Мезодерма задатка конечности получает инструкции от апикальной области и формирует все эти кости по очереди. Если апикальную область ранней стадии переместить на конечность, в которой уже сформировались некоторые кости, то он будет посылать инструкции по повторному формированию этих костей, так как ее внутренние часы находятся на более ранней стадии Нам пока неизвестен точный механизм этих часов, но, скорее всего, в нем участвуют регуляторные белки, которые на некоторое время связываются с определенными генами и запускают производство белков следующего вида. Теоретически нетрудно представить модель такого механизма. Регуляция по положению и развитие тела мухи Плодовая мушка оказалась в высшей степени полезной не только при изучении классических принципов наследственности, но и при исследованиях в области генетики развития. Будущую специализацию клеток определить легче всего по их местоположению. Глаза и рот должны развиваться на голове, конечности — в середине нижней (брюшной) части тела, крылья — на верхней части тела. Но как происходит изначальное деление на части тела? Если яйцеклетка однородна и практически симметрична, то, казалось бы, невозможно установить местонахождение будущих органов. Однако клетка развивается не отдельно сама по себе, а в окружении других клеток, которые и предоставляют ей пространственную информацию. Яйцеклетка дрозофилы развивается в яичнике мухи, в окружении 15 других клеток, называемых клетками-«нянями», от которых и зависят пространственные направления. Эти клетки связаны с развивающейся яйцеклеткой маленькими канальцами, через которые поступают вещества, необходимые для развития. Среди этих веществ могут быть специфические мРНК, которые начинают определять дифференциацию яйцеклетки (после того как она оплодотворена и стала зиготой). Принцип начальной дифференциации показан на рис. 11.3. Участвующие в этом процессе гены обозначены, как и обычно, по названиям мутаций, потому они имеют порой довольно забавные названия. Названия генов выделены курсивом; названия кодируемых ими белков начинаются с прописных букв и даны прямым шрифтом. Рис. 11.3. Начальные взаимодействия генов, определяющих пространственную ориентацию эмбриона плодовой мушки. В передний конец внедряется мРНК bicoid и синтезируемый на ее основе белок Bicoid включает транскрипцию гена hunchback. Это приводит к транскрипции генов головы и груди. В задний конец внедряется мРНК nanos u синтезируемый ею белок. Nanos подавляет экспрессию белка Hunchback. Как следствие включаются гены брюшка Решающим событием служит внедрение мРНК bicoid в тот конец, который позже станет передним (и где будет расположена голова), а также внедрение мРНК nanos в тот конец, который позже станет задним. Белки, транслируемые с каждой из этих мРНК, либо способствуют синтезу хотя бы одного из белков, либо подавляют синтез. Затем цепь синтеза белков идет по нарастающей, пока белки не образуют разные структуры. Как только начальное направление определено, включаются другие гены, которые делят эмбрион на сегменты, поскольку тело мухи членится на отдельные сегменты: 5 сегментов в области головы, 3 грудных сегмента и 11 в области брюшка. Задействованные при этом белки можно наблюдать при помощи подкрашивания их антител. Если развивающийся эмбрион погрузить в раствор антител, то антитела сконцентрируются там, где расположены соответствующие белки. Такие опыты показали, что сначала включается серия генов gap. Кодируемые ими белки активизируют серию генов pair-rule, которые делят эмбрион на 14 сегментов. Затем включаются гены segment-polarity, которые делят каждый сегмент на переднюю и заднюю части. После этого начинает действовать серия генов homeotic, которые определяют строение каждого сегмента с присущими ему признаками. Именно эти гены первыми были опознаны в мутантах необычной формы. Например, мутанты Antennapedia имеют на голове вместо усиков пару ног. Спаривая между собой различных го-меотических мутантов, Эд Льюис вместо обычных двукрылых вырастил четырехкрылых мушек. Формирование глаза мухи Одна из самых интересных серий событий с участием нескольких генов происходит при формировании глаза мушки дрозофилы. Сложный глаз насекомого состоит приблизительно из 800 элементов. Отдельный элемент, омматидий, состоит примерно из 20 клеток, расположенных в определенном порядке. Эти клетки образуют тонкий слой и взаимодействуют только со своими соседями; выше и ниже клеток нет. Обычно эти клетки становятся клетками хрусталика, но если они вступают в серию генетически запрограммированных взаимодействий друг с другом, то они специализируются в фоторецепторы, которые воспринимают свет и посылают в мозг сигналы. Дифференциация в каждом омматидии происходит в строго определенной последовательности (рис. 11.4). Сначала специализируется центральный фоторецептор, R8. Далее он вызывает специализацию своих соседей, R2 и R5. Потом они, в свою очередь, вызывают специализацию своих соседей с одной стороны, R3 и R4, и с другой стороны, R1 и R6. Под конец происходит специализация R7. Оставшиеся клетки группы становятся хрусталиком и другими структурами. Дифференциация начинается после того, как через слой клеток из передней в заднюю часть проходит волна, видимая, как заметное углубление. После прохождения волны клетки начинают делиться и затем останавливаются на ранней стадии клеточного цикла. Ориентация волны, а следовательно, и группы фоторецепторов, зависит от экспрессии гена wingless (wg), продуктом которого является белок, сигнализирующий о начале выработки серии белков, выполняющих специфические действия по дифференциации. Клетки за волной начинают дифференцироваться и выделять белок Hedgehog (Hh). Hh растворяется в недифференцированных клетках и стимулирует их производить белок Decapentaplegic (Dpp). Dpp растворяется в еще большем количестве передних клеток, заставляя их дифференцироваться и выделять Hh, который в свою очередь запускает очередной цикл производства Dpp. Так циклическое производство белков Hh и Dpp заставляет волну двигаться вперед.
Рис. 11.4. Восемь клеток, которые становятся фоторецепторами в омматидии глаза дрозофилы, специализируются в определенном порядке, который зависит от взаимодействия нескольких генов и кодируемых ими белков Чтобы понять, как специализируются следующие клетки, рассмотрим клетку R7, хотя она и специализируется в последнюю очередь. Мутация sevenless (sev) происходит в гене, который требуется для дифференциации R7. Клетка R7 отличается тем, что это рецептор ультрафиолетового света, и мутанты sevenless могут быть опознаны по тому, что они не воспринимают ультрафиолетовый свет. Белок Sevenless служит рецептором на поверхности клетки, которая станет R7, и, как лиганд, он связывается с белком Boss, который кодируется геном bride-of-sevenless (boss) и вырабатывается клеткой R8. Таким образом, белок Boss клетки R8 дает инструкции соседней клетке по превращению в специализированную клетку R7. Мы еще не знаем до конца, как именно запрограммированы все эти взаимодействия, но анализ некоторых мутантов вывил общую картину: одна из клеток вырабатывает определенные белки, которые взаимодействуют с белками соседних клеток, включая один набор генов и выключая другой. Если вернуться к вопросу, поставленному в начале главы, то придется ответить, что существуют разные механизмы развития. Одни гены кодируют белки, которые непосредственно связываются с другими генами, включая их или выключая. Другой класс генов кодирует белки, регулирующие трансляцию специфических мРНК в соответствующие им белки. Некоторые гены кодируют синтез белков, переходящих из одной клетки в другую и заставляющих их меняться или противодействующих переменам. Есть и такие гены, которые кодируют белки, взаимодействующие с другими белками на поверхности близлежащих клеток. Развитие эмбриона зависит от сложного переплетения разнообразных видов взаимодействий, среди которых мы упомянули лишь несколько самых распространенных. Глава двенадцатая
|
||||
|
|
Последнее изменение этой страницы: 2019-04-27; просмотров: 281; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.221.221.171 (0.007 с.) |