Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Глава 1 биологическая характеристика чавычи в связи со средой обитания и образом жизниСтр 1 из 5Следующая ⇒
ВВЕДЕНИЕ Рыбное хозяйство Российской Федерации в настоящее время набирает высокий темп развития. Появляются новые заводы по разведению рыб, которые обеспечивают работой население страны. Повышается спрос на рыбную продукцию отечественного производства не только в России, но и за рубежом. Контроль развития рыбной промышленности осуществляется на правительственном уровне. На данный момент принят Федеральный закон от 02.07.2013 № 148-ФЗ «Об аквакультуре (рыбоводстве) и о внесении изменений в отдельные законодательные акты Российской Федерации». Этот закон обеспечивает стабильное развитие аквакультуры, охрану и сохранение водных биоресурсов страны. Кроме того были приняты следующие документы Указ Президента Российской Федерации от 30 мая 2008 г. № 863 «О Федеральном агентстве по рыболовству» и постановление Правительства Российской Федерации от 11 июня 2008 г. № 444 «О Федеральном агентстве по рыболовству» (www.fish.gov.ru). Сильное антропогенное воздействие на окружающую среду пошатнуло состояние естественных популяций ценных промысловых рыб. Многие виды уже не могут восстановить свою численность естественным размножением. Поэтому искусственное воспроизводство является единственным путем сохранения таких видов. Оно кроме того обеспечит стабильные уловы менее редких видов. Основу искусственного воспроизводства составляет создание благоприятных условий созревания производителей, инкубации икры и развития молоди. Благодаря тому что все эти процессы находятся под тщательным контролем специалистов, выращенная молодь получается более жизнестойкой. А это значит, что и промысловый возврат от нее будет выше, чем от молоди естественного происхождения. Лососевые являются наиболее распространенной группой рыб, популяции которых поддерживаются с помощью искусственного воспроизводства. Искусственное воспроизводство дальневосточных лососей на Камчатке относительно невелико, по сравнению с естественным. Доля заводских рыб не превышает 0,5% от суммарных подходов производителей тех же видов. Камчатское лососеводство ориентированно в основном на выпуск молоди на жизнестойких стадиях. Однако большая часть ее гибнет и не возвращается на нерест в зрелом состоянии. Поэтому следует разрабатывать новые способы выращивания, улучшать биотехники разведения и создавать благоприятные условия для возврата как можно большего количества производителей на нерест. (http://oxotskoe.arktikfish.com).
Одним из наиболее ценных видов тихоокеанских лососей является чавыча. На Камчатке чавыча почти столетие служила наиболее излюбленным объектом коммерческого промысла, но, начиная с середины 1990-х гг., она уступила это место нерке, которая на международном рынке имеет более высокую коммерческую стоимость, чем чавыча. В бассейне р. Камчатки промысел чавычи ведут во всех населенных пунктах, расположенных вдоль русла реки, но до 90-95 % общего вылова всегда добывали в нижнем течении реки и в Камчатском заливе. Многие годы чавычу вылавливали специальными плавными сетями на 7-9 основных участках, рассредоточенных на протяжении первых 60 км вверх по течению. При этом в реке традиционно вылавливали до 80-90 % чавычи за путину. В последние годы чавычу добывают в основном как прилов при добыче нерки ставными неводами (Бугаев, 2007). Биотехника искусственного воспроизводства самого крупного и наиболее ценного по своим вкусовым качествам тихоокеанского лосося, чавычи все ещё недостаточно разработана. Чавыча обладает качествами, которые позволяют получать высокие результаты при искусственном воспроизводстве. К таким качествам относятся: приспособленность к различным местам обитания, высокая адаптивность к изменениям гидрохимических, гидрологических и метеорологических показателей, высокая скорость роста, короткий жизненный цикл, высокоразвитый хоминг, широкий спектр питания и высокая стоимость произведённой из неё продукции. Цель данной курсовой работы – изучить биологические особенности чавычи в связи с её искусственным разведением. Поставленная цель определила следующие задачи: - дать биологическую характеристику чавычи в связи со средой обитания и образом жизни: общая биология вида, основные периоды онтогенеза; - изучить влияние различных факторов среды на чавычи; - выбрать один из методов стимулирования половых продуктов у производителей чавычи и описать данный метод;
- описать биологические основы кормления чавычи, определить спектр питания исследуемого объекта в ранние периоды онтогенеза; - выбрать один из методов транспортировки чавычи на разных этапах онтогенеза; - подобрать подходящий район акклиматизации, описать водоем в котором можно акклиматизировать чавычу
Общая биология вида Чавыча по современной систематике занимает следующее положение: Домен Эукариоты – Eukaryota Царство Животные – Animalia Подцарство Многоклеточные – Eumetazoa Раздел Двусторонне симметричные – Bilateria Тип Хордовые – Chordata Подтип Позвоночные – Vertebrata Надкласс Челюстноротые – Gnathostomata Группа (ряд) Рыбы – Pisces Класс Костные рыбы – Osteichthyes Подкласс Лучеперые – Actinopterygii Надотряд Клюпеоидные – Clupeomorpha Отряд Лососеобразные – Salmoniformes Семейство Лососёвые – Salmonidae Род Тихоокеанские лососи – Oncorhynchus Вид Чавыча – Oncorhynchus tshawytscha (Walbaum, 1792) (Пономарев, 2012) (рис. 1).
Рисунок 1 – Чавыча Чавыча или Oncorhynchus tshawytscha (англ. - Chinook salmon, King salmon) относится к видам тихоокеанских лососей с длительным пресноводным и морским периодами жизни. Все рыбы погибают после первого нереста. Молодь чавычи проводит в пресных водах 1 -4 года, небольшая часть скатывается в первое лето жизни - сеголетками (0+ лет). В море чавыча живет 2 года, реже - 5 и, исключительно редко, - 1 и 6. Тело чавычи, как и у других лососевых, длинное, сжатое с боков, покрыто закругленной циклоидной чешуёй. Брюшные плавники, по сравнению с другими пресноводными рыбами, отодвинуты назад, расположены в задней части брюха довольно близко к анальному плавнику. Грудные расположены ниже к анальному плавнику. Спинных плавников два - настоящий и расположенный позади жировой плавничок, который характерен для всех лососевых. Плавники не имеют колючих лучей Бока чавычи на океанской фазе имеет яркий серебристый окрас. Спина и верх головы, как правило, зеленовато-голубого, красноватого или пурпурного цвета; брюхо белого цвета. Небольшую чавычу можно ошибочно принять за кижуча, но для чавычи как на океанской, так и на речной фазе характерны черные десны на нижней челюсти, а некрупные тёмные пятна покрывают не только ее верхнюю часть тела (до боковой линии) и хвостовой стебель, но и обе доли хвостового плавника. Кроме того, от других лососей чавыча отличается большим (больше 15) числом жаберных лучей. На океанской фазе самцы и самки чавычи и других проходных лососей не имеют особых отличий или же их трудно обнаружить с первого взгляда. Но когда приходит время нереста и лосось начинает заходить в реки, рыба сильно изменяется в форме и окрасе, при этом проявляются существенные различия между самцами и самками - так называемый половой диморфизм. С каждым днем миграции лососей вверх по реке к местам нереста рыба все больше меняется в цвете и претерпевает все больших физиологических изменений. Радикально меняется цвет тела - рыба стремительно теряет серебристую окраску, приобретая розовые, красные и бордовые тона, появляются красные и чёрные пятна. Тело Лосося также трансформируется: оно становится более высоким и угловатым, голова заметно удлиняется (особенно у самцов). Челюсти становятся крючкообразными: верхняя челюсть изгибается вниз, нижняя — вверх, а зубы более крупными. Эти метаморфозы особенно заметны у самцов, которые претерпевают значительно больше изменений, чем самки.
Брачный наряд чавычи выражен слабее, чем у других лососей, таких как чавыча, нерка или горбуша. Рыба, как и другие лососи, меняется в цвете, чернеет и приобретает ярко-красный, розовый или бордовый цвет. Однако, тело самца чавычи хоть и становится более высоким и угловатым, но характерный для многих других тихоокеанских лососей горб не образуется. Кроме того, челюсти самца никогда не приобретают той степени крючковатости, которая наблюдается у других видов, а у самок чавычи может быть вообще не заметна. Кроме того, самки чавычи, обычно, не полностью теряют свой серебристый окрас и часто имеют стальной отлив даже во время нереста (http://americanfishing.ru). Поскольку чавыча является проходной рыбой, ее главные места обитания - пресноводные реки, солоноватые устья и открытое море. В реках чавыча любит держатся на течении в довольно глубоких местах с галечным или гравийным дном. На таких участках нужно искать рыбу в местах с неровностями дна, на бровках, возле всевозможных укрытий и коряг. Устья являются для чавычи своеобразной транзитной зоной между морем и рекой. Замечено, что чем больше водной растительности в водах устья, тем лучше для лосося, поскольку в таких местах рыба находит укрытие от хищников, а также необходимую пищу, которая нужна рыбе для дальнейшего движения вверх по реке или в открытое море. В море чавыча может держаться как в прибрежных водах, так и мигрировать на тысячи километров вглубь Тихого океана. чавыча не является глубоководной рыбой и редко встречается на глубине более 35 метров. Чавыча предпочитает насыщенную кислородом довольно холодную воду. Температура воды в 10-14ºС является самой оптимальной для развития рыбы. Чавыча по образу жизни является типичной проходной рыбой. Появляется лосось на свет в быстротекущих пресноводных реках. Вылупившиеся мальки, как правило, проводят в реке, в зависимости от речной системы, от нескольких месяцев до двух лет, после чего скатываются в низовья и выходят в открытое море. В море чавыча проводит от 1 до 8 лет. В соленой воде рыба стремительно набирает массу: в среднем около 99 % своей массы Чавыча набирает именно в море. Когда приходит время нереста, взрослые особи чавычи сбиваются в стаи и начинают заходить в реки, двигаясь вверх по течению именно к тем местам, где рыба появилась на свет. Сроки нереста у лосося наследственные. Как только взрослая Чавыча заходит в реки на нерест, она практически перестает питаться и заканчивает свое половое созревание, продвигаясь вверх по реке к нерестилищам. На нерестилищах самка выбивает хвостом так называемые “нерестовые ямы” в крупной гальке и булыжниках, куда откладывает до 14 000 крупных икринок, после чего их оплодотворяет самец. В течении одной-двух недель после нереста чавыча погибает. На территории восточной России нерест чавычи начинается в июне и длится практически все лето. В Америке же выделяют как минимум 4 сезонных “хода” Чавычи: весенний, летний, осенний и зимний. Ход обозначается по времени, когда лосось заходит в устья рек для дальнейшей миграции к нерестилищам. Нередко даже в пределах одной речной системы наблюдается несколько разных “ходов” Чавычи с перерывами в несколько месяцев. Таким образом, часто возникает ситуация, когда в какой-то реке практически круглый год есть лосось (http://americanfishing.ru).
Еще в середине 1960-х гг. Б. Б. Вронским было высказано предположение о неоднородности этого вида лососей, в частности, о наличии в бассейне р. Камчатки обособленных сезонных рас - ранней и поздней чавычи. Предположение это основывалось на установленных отличиях в сроках и местах нереста двух различных по численности группировок чавычи. Впоследствии данные наблюдения были дополнены исследованиями специфики нерестовых бугров и развития икры. Группа поздней чавычи в бассейне р. Камчатки относительно малочисленна и ее промысловое значение в традиционных районах лова невелико. Однако ближе к верхнему течению реки (район р. Шапиной, пос. Долиновка, пос. Мильково) в уловах второстепенных рыбопользователей удельное значение ее существенно возрастает вследствие того, что она образует здесь более плотные преднерестовые скопления, в отличие от чавычи ранних подходов, распределяющейся выше по течению реки и в ее притоках. Ранняя и поздняя сезонные расы чавычи р. Камчатки различают прежде всего по срокам хода и нереста. В свою очередь, они имеют различия в нерестовых стациях, возрастной структуре, длине и массе тела, экологии размножения, морфометрических и других характеристиках. Необходимо отметить, что особи поздней чавычи отличаются от ранней в той же стадии зрелости более интенсивной красной окраской. В то время как у нерестящихся или поздненерестящихся производителей ранней чавычи преобладают красно-буроватые тона, у производителей поздней расы основной тон составляет более светлый алый цвет, несколько темнеющий к концу нерестового периода. В р. Камчатку отдельные экземпляры чавычи заходит в середине мая. Интенсивный ход начинается в первой декаде нюня и продолжается до начала июля. В уловах чавыча стабильно встречается до начала середины августа, а позже (до середины сентября) - отмечаются только отдельные экземпляры. Чавыча заходит в реку в основном с гонадами в III стадии зрелости, но особи, мигрирующие позднее, имеют более развитые половые продукты, а у некоторых из них уже при заходе в реку могу т быть заметны брачные изменения, выражающиеся в изменении окраски, т. к. преднерестовые изменения экстерьера у чавычи относительно невелики.
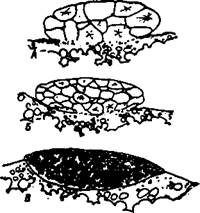
В отличие от других видов лососей (кроме горбуши), чавыча нерестится только на речных участках - в главных руслах рек и речных протоках. Почти все речные участки, где располагаются нерестилища чавычи, имеют ярко выраженный предгорно-горный характер. Как правило, подавляющая масса чавыч и нерестится в условиях типично речного режима. Она охотно размещается в зонах, где проявляется воздействие подрусловых потоков, а мест, где выходят фунтовые воды, избегает. В соответствии с этим нерестилища чавычи обычно расположены в русле реки на относительно быстром течении от 0,6-0,8 до 1,5-1,8 м/сек, а нередко и при скорости 0,3-0,5 м/сек. Нижний предел - 0,2 м/сек, верхний - 2-3 м/сек и даже более. Таким образом, эта крупная сильная рыба может избирать для нереста такие участки, где ей приходится противостоять чрезвычайно большому напору воды (лососи других видов нереститься в таких условиях не могут). Для поздней чавычи в конце нерестового периода характерно своеобразное поведение - производители над гнездами периодически выбрасываются из воды, совершая в воздухе кульбиты три-четыре раза подряд. Такие действия повторяются с различными интервалами - от 15-20 минут до нескольких часов. Аналогичное поведение свойственно также и сносимым по течению отнерестившимся рыбам, но оно никогда не наблюдалось у ранней чавычи. Основная масса молоди чавычи выходит из гнезд в верховьях р. Камчатки в конце апреля - первой половине мая, в период начала повышения уровня и температуры воды, связанного со снеготаянием. В период паводка, когда заливаются обширные участки поймы, молодь чавычи частично сносится течением, частично расселяется по заливаемым участкам, находя укрытия в местах со слабым течением вдоль береговой линии и в руслах проток за крупными камнями, неровностями рельефа дна и замытыми стволами и корнями деревьев. В верховьях р. Камчатки в середине-конце мая начинается массовый скат сеголетков чавычи и нерки. В это время среди десятков и сотен мальков указанных видов, сеголетки чавычи встречаются гораздо реже единицами и десятками. Эта молодь, скорее всего, вновь задержится в русле или протоке р. Камчатки в каком-либо защищенном от быстрого течения месте (Бугаев, 2007). Эмбриональное развитие 1. Обводнение икры, образование зародышевого диска. По данным Риддла (Riddle, 1917), развивающаяся икра чавычи имеет диаметр 7–9 мм (среднее 7,5 мм) и вес 260—330 мг. Прескотт (Prescott, 1955) сообщает, что вес икринок в результате обводнения возрастает на 15,5%, а объем — на 16,5%. В опытах с тяжелой водой (D2O) он убедительно доказал образование перивителлиновой жидкости за счет поступления в яйцо воды из внешней среды. Вес икринок разных самок из р. Паратунки после набухания увеличивался от 4,7 до 13,6%, их диаметр равнялся 7,5— 7,7 мм, средний вес 227—258 мг (вес отдельных икринок достигал 267 мг). Было замечено, что меньше воды воспринимала та икра, которая дольше задерживалась в теле самки. Во время набухания оболочка икры становится полупрозрачной, плотной, эластичной. Ее толщина, по данным Риддла, 64,7 мк. У набухших икринок камчатской чавычи толщина оболочек колебалась от 38,0 до 57,0 мк (53,75±0,78). Набухшие икринки покрывались белесым глянцеватым налетом, дающим в отраженном свете характерный перламутровый блеск. Микропилярное поле, имеющее диаметр немного более 1 мм, лишено такого налета. 2 этап. Дробление зародышевого диска. Первая бороздка дробления формировалась примерно через 4,5 градусо-дня после осеменения. Последующие дробления наступали через 5,2–5,5, 6.3–6,6, 7,4–7.7, 8,5–8,8, 9,7–10.0 и 10,8–11,2 градусо-дней. Борозды дробления располагались, как и у других лососей. По Риддлу (1917), при пятом дроблении борозды проходят в экваториальной плоскости и образуют зачаток из двух слоев клеток. До стадии 64 бластомеров зародышевый диск сохраняет немного вытянутую форму, а со стадии 128 бластомеров становится округлым. На этой стадии между нижележащими бластомерами образуются межклеточные промежутки, которые в последующем, по мнению Риддла, дают полость между бластодермой и перибластом (рис. 2). На нашем материале в экваториальной плоскости проходили борозды 4-го дробления, образуя 2 слоя клеток (рис. 3). Первая борозда проходила глубже других и ее расположение можно было выявить до образования 64 бластомеров. После стадии 128 клеток (в инкубаторе в возрасте 1 сут. 22 час.) бластодиск приобретал округлую форму с диаметром около 1 мм. До стадии 128 бластомеров чавыча развивается при температуре не ниже 4,4–5,8°. Верхний температурный порог лежит в пределах 14,2–15,5° (Combs, Burrows, 1957; Combs, 1965).
Рис. 2. Стадни дробления (А и Б) н бластула (В) чавычи (по Риддлу, 1917)
В возрасте 3 сут. (21 градусо-день) бласюдиск имел диаметр 1,0–1,2 мм и высоту 0,45–0,49 мм. Диаметр клеток поверхностного слоя был равен 75–80 мк. Позже более мелкие и плотно прилегающие друг к другу клетки покрывали массу свободно расположенных клеток. Снизу бластодиск подстилал нерибласт; при температуре 7,5–12° начало его формирования отмечено через 44 час. после осеменения.
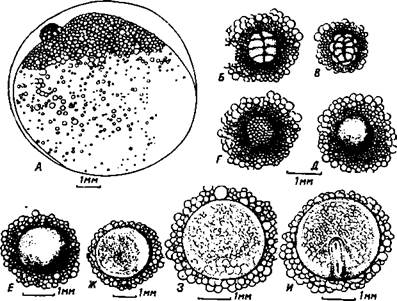
Рис. 3. Развитие чавычи от стадий дробления до образования головы и туловища эмбриона А – вид живого яйца сбоку в проходящем свете в начале дробления. Б-Л – бластодиски п зародыши с участком желточного мешка на разных стадиях развития. Б – 8 бластомеров. В – 16 бластомеров. Г – крупноклеточная морула. Д – морула мелких клеток. Е – бластула. Ж – начало гаструляции. З – стадия «зародышевого узелка». И – стадия «зародышевой полоски». К – начало сегментации туловищной мезодермы, длина зародыша 2,4 мм. Л – зародыш длиной 3,3 мм с 21 сомитом. М – фиксированная икринка без оболочки с эмбрионом, имеющим 25–26 сомитов
При 4° тысяча икринок в возрасте 4 сут. потребляла за 1 час 1,41 мг кислорода (среднее по двум определениям). Эта величина меньше по сравнению с потреблением кислорода во время набухания икры. Позже интенсивность обмена постепенно возрастает (табл. 1). Таблица 1 – Потребление кислорода эмбрионами и личинками чавычи
Продолжение таблицы 1
Примечание. Опыты с эмбрионами в возрасте [66 сут. и более проведены в 1956 г., остальные—в 1957 г., когда определения выполнялись Л. Е. Грачевым. 3 этап. Бластула. В возрасте 5 сут. 9 час. зародышевый диск имел вид, сходный с описанным Риддлом через 101 час после оплодотворения (рис. 2, В). Его диаметр увеличился до 1,35–1,5 мм. Диаметр клеток покровного слоя бластулы уменьшился до 40 мк (рис. 3, Е; 26–28 градусо-дней). Со временем бластодиск расширялся и уплотнялся. В первой стадии этап длился около 5,5 сут., во второй – немного более суток. 4 этап. Гаструляция. При температуре 3,7-4,1° признаки гаструляции отмечены в начале 11 сут., при 15,5-16,5° – в середине 4 сут. Зародышевый диск имел диаметр 1,7-2,1 мм. Диаметр клеток поверхностного слоя уменьшился до 20-25 мк. Через утоньшенную центральную часть диска просматривались лежащие снизу мелкие жировые капли. С каудальной стороны закладывающегося зародыша краевой участок диска начал утолщаться (рис. 3, Ж). С увеличением диаметра бластодиска до 2,5—3,2 мм (68 градусо-дней) образовался «краевой узелок» (рис. 3, З). Границы эмбрионального щита вышли за зону скопления мелких жировых капель. В речных аппаратах в возрасте 5 сут. 5 час. (около 75 градусо-дней) образовалась зародышевая полоска длиной 1,3-1,4 мм (рис. 3, И). С началом гаструляции икринки стали очень чувствительными к толчкам. 5 этап. Формирование головы и туловища зародыша. В икринках первой серии сегменты появились в возрасте 19-20 сут. (80-84 градусо-дня), во второй — в начале 6-х сут. К этому времени диаметр бластодиска достигал 3,5-4 мм, а длина зародышей – 2-2,2 мм. Некоторые эмбрионы в речных аппаратах в возрасте 5 сут. 17 час. имели по 3-4 сегмента; у них наметилась дифференцировка отделов головного мозга (рис. 3, К). В инкубаторе через 2 сут. после начала сегментации эмбрионы достигали длины 3,2 мм. У них сформировалось по 7-8 сомитов, обозначились зачатки глаз. Через сутки, на стадии 12-13 сомитов, появились зачатки хрусталиков и слуховые пузырьки. Когда у эмбрионов насчитывалось 20-22 сегмента (около 100 градусо- дней), головной отдел составлял 34,5% их длины (рис. 3, Л). В возрасте 25 сут. (104 градусо-дня) бластодерма охватывала до половины сферы желточного мешка. Зародыши достигали длины 3,3—3,6 мм, у них насчитывалось 25—26 сегментов. Образовалась перикардиальная полость, в которой размещался зачаток сердца. Основная масса жировых капель сосредоточивалась вблизи тела эмбриона и лишь немногие более мелкие находились за пределами зоны обрастания (рис. 3, М). К 34 сут. (138 градусо-дней) бластодерма покрывала 2/3 поверхности желточного мешка или немного большую часть. В икре из речных аппаратов сходная стадия наблюдалась в возрасте 8 сут. (115 градусо-дней). Эмбрионы достигли длины 4,1-4,4 мм и у них насчитывалось 35-37 сегментов (рис. 4, А). Образовалась небольшая хвостовая почка. Размер головного отдела сократился до 28% длины зародыша. Слуховые капсулы от глаз были удалены на расстояние, почти равное ширине 10 сегментов. Относительно небольшие глаза имели диаметр около 0,4 мм. Обозначились обонятельные плакоды. Четко выявились отделы головного мозга и энцефаломеры. Когда бластодерма покрыла большую половину желточного мешка, чувствительность икринок к толчкам на некоторое время снизилась. 6 этап. Обособление задней части тела зародыша от поверхности желточного мешка. В конце 36 – начале 37 сут. (145-150 градусо-дней) зародыши достигли длины 4,9-5,2 мм и у них сформировалось 45-48 сегментов. Сохранялась лишь небольшая желточная пробка. Продолжала дифференцироваться хвостовая почка. Появились зачатки жаберных крышек. В этом состоянии наблюдались слабые сокращения миотомов. Сердечная трубка приняла изогнутую форму и у эмбрионов с большим числом миотомов волнообразно сокращалась. При развитии в речных аппаратах пульсация сердца отмечена в возрасте 9 сут., по прошествии всего лишь 129 градусо-дней. В этом возрасте чувствительность икры к толчкам резко возрастала.
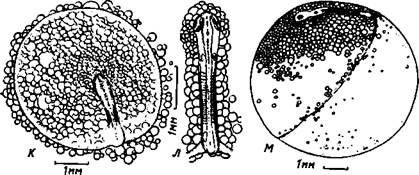
Рис. 4. Чавыча на стадиях завершения процесса эпиболии и начала развития кровеносной системы. А – стадия, близкая к завершению процесса эпиболии, длина эмбриона 4,1 мм. Б – начало кровообращения, длина эмбриона 6,6 мм. В – стадия появления задних кардинальных вен, длина эмбриона 8,5 мм. Г – стадия появления в глазах меланина, длина эмбриона 9,8 мм
В мелких икринках первой серии процесс эпиболии завершался у эмбрионов длиной 5,5-6,0 мм на стадии 48-49 сегментов (развитие с 5/VIII по 14/IX 1956 г., около 160 градусо-дней). В это время длина головы составляла 23% длины эмбрионов. От желточного мешочка отделилась хвостовая почка, равная ширине семи туловищных миотомов. Ниже 43—44 сомитов сформировалось анальное отверстие. Появился зачаток непарной плавниковой складки. У основной массы зародышей эпиболия завершалась при 53-56 сегментах, а в крупных икринках – после образования 58 и 59 сегментов. При более высокой температуре у отдельных икринок обрастание желточного мешка бластодермой продолжалось до стадии 63—64 сегментов. Таким образом, в зависимости от качества икры (ее размеров) и условий инкубации процесс эпиболии заканчивается на разных стадиях морфологической дифференцировки. Имеется указание на завершение этого процесса у эмбрионов североамериканской чавычи длиной 4 мм, имевших 38 сегментов (Riddle, 1917). Такое ускорение эпиболии, возможно, свидетельствует о генетических особенностях материала, который был в распоряжении Риддла. Во время завершения эпиболии икра очень чувствительна к толчкам. 7 этап. Развитие подкишечно-желточной системы кровообращения. В условиях низкой температуры циркуляция крови у разных эмбрионов обнаруживалась через 43-47 сут. после оплодотворения (172-187 градусо-дней), когда они достигали длины 7-7,5 мм и имели по 64-66 сегментов. К этому времени хвост, в котором насчитывалось 21-22 сегмента, и задний отдел туловища обособились от желточного мешка. Голова по-прежнему плотно прилегала к желточному мешку, а ее длина продолжала уменьшаться. Появились зачатки жаберных крышек. Начали формироваться обонятельные ямки. Слуховые пузырьки заметно увеличились; в них дифференцировались полукружные каналы и были видны многочисленные мелкие отолитики. От 15 или 17-го миотома вдоль спины тянулась узкая плавниковая складка. Значительной длины достигла преанальная складка. Заложились грудные плавнички. Позади них просматривались клубочки пронефроса. Наметилось обособление желудка. В начале этапа форменных элементов в сосудах мало, эритроциты окрашены слабо, кровь течет медленно и омывает лишь два сегмента хвостового отдела. Схема кровообращения сходна с описанной у кеты (рис. 4, Б). У эмбрионов, развивающихся в речных аппаратах, кровообращение начиналось при 63-64 сегментах (10 1/4 сут., 147 градусо- дней). У некоторых из них оставалась небольшая желточная пробка. Эмбрионы периодически наклонялись то на правый, то на левый бок и медленно поводили хвостами. При температуре около 14,5° этап длился менее суток, при 3,5-3,7° – около 4 сут. 8 этап. Возникновение кардинальных вен и смешанного подкишечно-желточного и печеночно-желточного кровообращения. В первой серии между 49 и 52 сут. (195-207 градусо-дней) кровеносные сосуды эмбрионов обильно наполнились форменными элементами. Зародыши достигли длины 7,8-8,5 мм (рис. 4, В). Число сегментов в хвосте увеличилось до 35. Голова зародышей начала отделяться от желточного мешка. Расстояние между глазами и слуховыми капсулами сократилось до ширины 5 туловищных миотомов. Углубилась ротовая воронка. Появились зачатки нижних челюстей. Расширившаяся непарная плавниковая складка достигла 4–5-го туловищного сегментов. Кровью снабжалось 18—20 сегментов хвоста. Сформировались задние кардинальные вены, образовались кювьеровы протоки. Подкишечно-желточная вена опустилась ниже скопления жировых капель. На желточном мешке появилось 5—6 тонких капилляров. При развитии в речных аппаратах сходное состояние кровеносной системы отмечено в конце 11 сут. — в начале 12 сут. (160— 165 градусо-дней), когда зародыши имели длину 7—7,3 мм. У зародышей 1-й серии в возрасте 55 сут. (218 градусо-дней), а во 2-й — в 14 сут. (209 градусо-дней) начиналось печеночное кровообращение (по времени немного позже, чем у нерки и кижуча), появился ток крови в первой паре жаберных артерий, образовались первые сегментальные сосуды. В этом возрасте в глазах появился меланин. На рис. 4, Г изображен зародыш через 2 дня после появления в глазах меланина. Такие эмбрионы имели длину 9,5—9,9 мм и 77—78 миотомов. Их хвостовой отдел составлял 30—30,5% всей длины. В почечных клубочках и мочеточнике появились кристаллы, что указывает на начало функционирования органа. Сегментальные сосуды образовались по всему туловищу и в первых сегментах хвоста. Сетью капилляров покрылась примерно половина поверхности желточного мешка. В речных аппаратах в начале ригментации глаз зародыши имели длину 8,6—9,1 мм. У них сформировался 80—81 миотом, то есть сегментация закончилась. Кровью снабжались две пары жа-бер. Начала развиваться капиллярная сеть на дорзальной поверхности головы. При температуре 13,5—14° сердце сокращалось 50—51 раз в минуту. 9 этап. Формирование печеночно-желточной системы кровообращения. При температуре около 4° течение крови из печени отмечено на 62—63-е-сут. Эмбрионы достигли длины 10,3—11,8 мм и имели 78—80 миотомов (из них хвостовых 37—38), сегментация закончилась. Голова стала массивной. Слуховые капсулы увеличились, их основания расширились, образовались полукружные каналы и крупные отолиты. Кончик хорды начал изгибаться кверху и хвост утратил симметричность. Основания грудных плавников приняли наклонное положение. Наполнились кровью 1 или 2 пары жаберных артерий. Кровью не снабжалось только 6—8 последних хвостовых миотомов. Капиллярами покрылся весь желточный мешок, исключая небольшой участок ниже головы. В течение этого этапа быстро увеличивалась длина хвоста. У эмбрионов из речных аппаратов переход к печеночно-желточному кровообращению произошел на 18-е сут. (260 граду со-дней), когда они имели длину 10—11,5 мм (рис. 5, А). Кровью снабжались уже 3 пары жаберных дуг, наблюдался слабый ток крови в гиоидном сосуде. Появились капилляры на верхней поверхности головы. В большинстве хвостовых миотомов образовались сегментальные сосуды. Уплотнилась сеть капилляров на желточном мешке (на 1 мм2 его поверхности длина капилляров достигала примерно 4 мм). 10 этап. Формирование верхних и нижних конусов миотомов. Зачатки верхних и нижних конусов миотомов у зародышей первой серии появились на 68—70-е сут. (270—276 градусо-дней), эмбрионы имели длину 12—13 мм. Их голова стала выше, но продолжала оставаться относительно короткой. Хвостовой отдел составлял 32— 33% общей длины. Непарная плавниковая складка достигла наибольшей длины и заметно расширилась. Наметились зачатки непарных плавников. Кровью снабжались три пары жабер. На дорзальной поверхности головы развилась густая сеть капилляров. Сосудами покрылась вся поверхность желточного мешка. Образовалась нижняя приносящая желточная вена. Выявились первые железки вылупления. На поверхности головы, вблизи крупных сосудов и на выстилке брюшной полости появились первые меланофоры. На рис. 5, Б изображен эмбрион через несколько дней после начала этапа, развивавшийся в инкубаторе 75 сут.
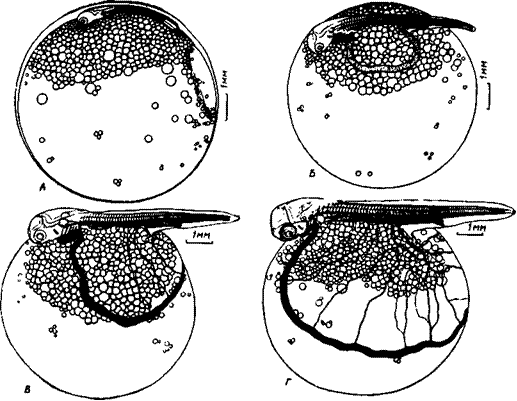
Рис. 5. Чавыча па стадиях перехода к печеночно-желточному кровообращению п дифференциации миотомов. А — стадия завершения сегментации хвостовой мезодермы и появления кровотока в гиоидных дугах, длина эмбриона 11,4 мм. Б — стадия начала формирования верхних н нижних конусов миотомов, закладки непарных плавников и появления меланофоров, длина эмбриона 12,9 мм. В — стадия появления первых лепидотрихий в хвостовом плавнике и липофоров в покровах головы, длина эмбриона 14,5 мм.
У эмбрионов второй серии этап начался в возрасте 21 сут. (302 градусо-дня). Они имели длину около 13,5 мм и отличались более длинным хвостом (до 37% общей длины). Грудные плавники приобрели подвижность. В глазах появился гуанин, но еще отсутствовали меланофоры на голове и теле. Сегментальные сосуды сформировались во всех миотомах и в первых миотомах начали разветвляться. Через 3 сут. такие зародыши достигли длины 14—15 мм (рис. 5, В; 344 градусо-дня). Их слуховые капсулы были максимально сближены с глазами, которые стали черными. Жаберные крышки прикрыли первую пару жабер. Нижние челюсти удлинились и, так же как жаберные крышки, приобрели подвижность. Верхние и нижние конусы развились на всех миотомах. Хвостовой отдел превысил 38% длины эмбрионов. Последние хвостовые миотомы распадались, и их число сократилось до 34—35. Сформировались три гипуралии. В хвостовом плавнике заложились первые лучи. Началось окостенение элементов плечевого пояса. Грудные плавники стали снабжаться кровью. Покрылась густой сетью капилляров верхняя поверхность головы. Кровеносные сосуды появились в основании лопасти хвостового плавника. На 1 мм2 поверхности желточного мешка длина капилляров превысила 7 мм. На данной стадии в покровах головы появляются малиново-красные липофоры (на рис. 5, В они показаны черными мелкими точками). Эмбрионы большинство времени оставались неподвижными, лишь изредка поворачивались с боку на бок. Периодически они совершали плавничками 10—15 взмахов в мин. По кишке время от времени проходили волны слабых перистальтических движений. У зародышей, развивавшихся в инкубаторе, мускульные почки в основании спинного плавника появились на 76—79-е сут. (300— 310 градусо-днсй). По сравнению с зародышами, развивавшимися в аппаратах с речной водой, они имели относительно укороченный хвост, длина которого составляла около 35% всей длины. Эти эмбрионы отличались также слабее развитой сосудистой сетью, а меланофоров имели больше. В возрасте 83 сут. (326 градусо-дней) зародыши достигли длины 13,5—14,7 мм. У них заложились брюшные плавники. Грудные плавники приобрели подвижность. В просвете кишки был виден желто-зеленоватый пигмент. Повысилась вязкость желтка.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 405; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.86.134 (0.07 с.) |