Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Хвостатое ядро Чечевицеобразное ядроСодержание книги
Поиск на нашем сайте
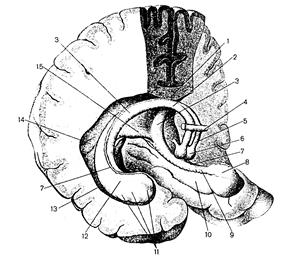
В толще белого вещества каждого полушария большого мозга имеются скопления серого вещества, образующего отдельно лежащие ядра (рис.7). Эти ядра залегают ближе к основанию мозга и называются базальными (подкорковыми, центральными). К ним относятся: 1) полосатое тело, которое у низших позвоночных составляет преобладающую массу полушарий; 2) ограда; 3) миндалевидное тело. Рассмотрим строение полосатого тела (corpus striatum), которое на разрезах мозга имеет вид чередующихся полос серого и белого вещества. Наиболее медиально и впереди находится: а) хвостатое ядро, расположенное латеральнее и выше таламуса, будучи отделенным от него коленом внутренней капсулы. Ядро имеет головку, залегающую в лобной доле, выступающую в передний рог бокового желудочка и примыкающую к переднему продырявленному веществу. Тело хвостатого ядра лежит под теменной долей, ограничивая с латеральной стороны центральную часть бокового желудочка. Хвост ядра участвует в образовании крыши нижнего рога бокового желудочка и достигает миндалевидного тела, лежащего в переднемедиальных отделах височной доли (кзади от переднего продырявленного вещества); б) чечевицеобразное ядро- расположено латерально от хвостатого ядра. Прослойка белого вещества – внутренняя капсула – отделяет чечевицеобразное ядро от хвостатого ядра и от таламуса.
Рис.7. Фронтальный разрез головного мозга на уровне сосцевидных тел. 1–сосудистое сплетение бокового желудочка (центральная часть), 2–таламус, 3–внутренняя капсула, 4–кора островка, 5–ограда, 6–миндалевидное тело, 7–зрительный тракт, 8–сосцевидное тело, 9–бледный шар, 10–скорлупа, 11–свод мозга, 12–хвостатое ядро, 13–мозолистое тело.
На фронтальном разрезе головного мозга чечевицеобразное ядро также имеет форму треугольника, вершина которого обращена в медиальную, а основание в латеральную сторону (рис.7). Две параллельные вертикальные прослойки белого вещества делят чечевицеобразное ядро на три части. Наиболее латерально лежит более темная скорлупа, медиальнее находится «бледный шар», состоящий из двух пластинок: медиальной и латеральной. Хвостатое ядро и скорлупа относятся к филогенетически более новым образованиям, бледный шар – к более старым. Ядра полосатого тела образуют стриопаллидарную систему, которая, в свою очередь, относится к экстрапирамидной системе, участвующей в управлении движениями, регуляции мышечного тонуса (рис.).
Рис.8. Горизонтальный разрез головного мозга. Базальные ядра.
Тонкая вертикально расположенная ограда, залегающая в белом веществе полушария сбоку от скорлупы, отделена от скорлупы наружной капсулой, от коры островка – самой наружной капсулой. Хвостатое ядро и скорлупа получают нисходящие связи преимущественно от экстрапирамидной коры через подмозолистый пучок. Другие поля коры большого мозга также посылают большое количество аксонов к хвостатому ядру и скорлупе. Основная часть аксонов хвостатого ядра и скорлупы идет к бледному шару, отсюда - к таламусу и только от него - к сенсорным полям. Следовательно, между этими образованиями имеется замкнутый круг связей. Хвостатое ядро и скорлупа имеют также функциональные связи со структурами, лежащими вне этого круга: с черной субстанцией, красным ядром, люисовым телом (субталамическое ядро), ядрами преддверия, мозжечком, гамма- клетками спинного мозга. Обилие и характер связей хвостатого ядра и скорлупы свидетельствуют об их участии в интегративных процессах, организации и регуляции движений, регуляции работы вегетативных органов. Медиальные ядра таламуса имеют прямые связи с хвостатым ядром, свидетельством чего служит реакция его нейронов, наступающая через 2-4 мс после раздражения таламуса. Реакцию нейронов хвостатого ядра вызывают раздражения кожи, световые, звуковые стимулы.
При недостатке дофамина в хвостатом ядре (например, при дисфункции черного вещества) бледный шар растормаживается, активизирует спинно-стволовые системы, что приводит к двигательным нарушениям в виде ригидности мышц. Хвостатое ядро и бледный шар принимают участие в таких интегративных процессах, как условнорефлекторная деятельность, двигательная активность. Это выявляется при стимуляции хвостатого ядра, скорлупы и бледного шара, деструкции и при регистрации электрической активности. Прямое раздражение некоторых зон хвостатого ядра вызывает поворот головы в сторону, противоположную раздражаемому полушарию, животное начинает двигаться по кругу, т.е. возникает так называемая циркуляторная реакция. У человека стимуляция хвостатого ядра во время нейрохирургической операции нарушает речевой контакт с больным: если больной что-то говорил, то он замолкает, а после прекращения раздражения не помнит, что к нему обращались. В случаях травм головного мозга с раздражением головки хвостатого ядра у больных отмечается ретро-, антеро-, ретроантероградная амнезии. Раздражение хвостатого ядра может полностью предотвратить восприятие болевых, зрительных, слуховых и других видов стимуляции. Раздражение вентральной области хвостатого ядра снижает, а дорсальной - повышает слюноотделение. В случае повреждения хвостатого ядра наблюдаются существенные расстройства высшей нервной деятельности, затруднение ориентации в пространстве, нарушение памяти, замедление роста организма. После двустороннего повреждения хвостатого ядра условные рефлексы исчезают на длительный срок, выработка новых рефлексов затрудняется, общее поведение отличается застойностью, инертностью, трудностью переключений. При воздействиях на хвостатое ядро, помимо нарушений высшей нервной деятельности, отмечаются расстройства движения. Многие авторы отмечают, что у разных животных при двустороннем повреждении полосатого тела появляется безудержное стремление двигаться вперед, при одностороннем - возникают манежные движения. Для скорлупы характерно участие в организации пищевого поведения: пищепоиска, пищенаправленности, пищезахвата и пищеварения- ряд трофических нарушений кожи, внутренних органов возникает при нарушениях функции скорлупы. Раздражения скорлупы приводят к изменениям дыхания, слюноотделения. Как упоминалось ранее, раздражение хвостатого ядра тормозит условный рефлекс на всех этапах его реализации. В то же время раздражение хвостатого ядра препятствует угашению условного рефлекса, т.е. развитию торможения; животное перестает воспринимать новую обстановку. Учитывая, что стимуляция хвостатого ядра приводит к торможению условного рефлекса, следовало бы ожидать, что разрушение хвостатого ядра вызывает облегчение условнорефлекторной деятельности. Но оказалось, что разрушение хвостатого ядра тоже приводит к торможению условнорефлекторной деятельности. Видимо, функция хвостатого ядра не является просто тормозной, а заключается в корреляции и интеграции процессов оперативной памяти. Это подтверждается также тем, что на нейронах хвостатого ядра конвергирует информация различных сенсорных систем, так как большая часть этих нейронов полисенсорна.
Бледный шар имеет преимущественно крупные нейроны Гольджи 1 типа. Связи бледного шара с таламусом, скорлупой, хвостатым ядром, средним мозгом, гипоталамусом, соматосенсорной системой свидетельствуют об его участии в организации простых и сложных форм поведения. Раздражение бледного шара с помощью вживленных электродов вызывает сокращение мышц конечностей, активацию или торможение- гамма мотонейронов спинного мозга. Стимуляция бледного шара в отличие от стимуляции хвостатого ядра не вызывает торможения, а провоцирует ориентировочную реакцию, движения конечностей, пищевое поведение (обнюхивание, жевание, глотание и т.д.). Повреждение бледного шара вызывает у людей гипомимию, маскообразность лица, тремор головы, конечностей (причем этот тремор исчезает в покое, во сне и усиливается при движениях), монотонность речи. При повреждении бледного шара наблюдается миоклония - быстрые подергивания мышц отдельных групп или отдельных мышц рук, спины, лица. В первые часы после повреждения бледного шара в остром опыте на животных резко снижалась двигательная активность, движения характеризовались дискоординацией, отмечалось наличие незавершенных дискоординаций, незавершенных движений, при сидении – поникшая поза. Начав движение, животное долго не могло остановиться. У человека с дисфункцией бледного шара затруднено начало движений, исчезают вспомогательные и реактивные движения при вставании, нарушаются содружественные движения рук при ходьбе, появляется симптом пропульсии: длительная подготовка к движению, затем быстрое движение и остановка. Такие циклы у больных повторяются многократно. Ограда содержит полиморфные нейроны разных типов. Она образует связи преимущественно с корой большого мозга. Глубокая локализация и малые размеры ограды представляют определенные трудности для ее физиологического исследования. Это ядро имеет форму узкой полоски серого вещества, расположенного под корой большого мозга в глубине белого вещества. Стимуляция ограды вызывает ориентировочную реакцию, поворот головы в сторону раздражения, жевательные, глотательные, иногда рвотные движения. Раздражение ограды тормозит условный рефлекс на свет, мало сказываются на условном рефлексе на звук. Стимуляция ограды во время еды тормозит процесс поедания пищи. Известно, что толщина ограды левого полушария у человека несколько больше, чем правого; при повреждении ограды правого полушария наблюдается расстройство речи.
Таким образом, базальные ядра головного мозга являются интегративними центрами организации моторики, эмоций, высшей нервной деятельности, причем каждая из этих функций может быть усилена или заторможена активацией отдельных образований базальных ядер. Миндалевидное тело залегает в белом веществе височной доли полушария, примерно на 1,5–2 см кзади от височного полюса. Миндалевидное тело (corpus amygdoloideum), миндалина - подкорковая структура лимбической системы, расположенная в глубине височной доли мозга. Нейроны миндалины разнообразны по форме, функциям и нейрохимическим процессам в них. Функции миндалины связаны с обеспечением оборонительного поведения, вегетативными, двигательными, эмоциональными реакциями, мотивацией условнорефлекторного поведения. Электрическая активность миндалин характеризуется разноамплитудными и разночастотными колебаниями. Фоновые ритмы могут коррелировать с ритмом дыхания, сердечных сокращений. Миндалины реагируют многими своими ядрами на зрительные, слуховые, интероцептивные, обонятельные, кожные раздражения, причем все эти раздражения вызывают изменения активности любого из ядер миндалины, т.е. ядра миндалины полисенсорны. Реакция ядра на внешние раздражения длится, как правило, до 85 мс, т.е. значительно меньше, чем реакция на подобные же раздражения новой коры. Нейроны имеют хорошо выраженную спонтанную активность, которая может быть усилена или заторможена сенсорными раздражениями. Многие нейроны полимодальны и полисенсорны и активируются синхронно с тета-ритмом. Раздражение ядер миндалевидного тела создает выраженный парасимпатический эффект на деятельность сердечно-сосудистой, дыхательной систем, приводит к понижению (редко к повышению) кровяного давления, урежению сердечного ритма, нарушению проведения возбуждения по проводящей системе сердца, возникновению аритмии и экстрасистолии. При этом сосудистый тонус может не изменяться. Урежение ритма сокращений сердца при воздействии на миндалины отличается длительным скрытым периодом и имеет длительное последствие Раздражение ядер миндалины вызывает угнетение дыхания, иногда кашлевую реакцию. При искусственной активации миндалины появляются реакции принюхивания облизывания, жевания, глотания, саливации, изменения перистальтики тонкой кишки, причем эффекты наступают с большим латентным периодом (до 30-45 с после раздражения). Стимуляция миндалин на фоне активных сокращений желудка или кишечника тормозит эти сокращения. Разнообразные эффекты раздражения миндалин обусловлены их связью с гипоталамусом, который регулирует работу внутренних органов. Повреждение миндалины у животных снижает адекватную подготовку автономной нервной системы к организации и реализации поведенческих реакций, приводит к гиперсексуальности, исчезновению страха, успокоению, неспособности к ярости и агрессии. Животные становятся доверчивыми. Например, обезьяны с поврежденной миндалиной спокойно подходят к гадюке, вызывавшей ранее у них ужас, бегство. Видимо, в случае повреждения миндалин исчезают некоторые врожденные безусловные рефлексы, реализующие память об опасности.
К белому веществу полушария относятся внутренняя капсула и волокна, имеющие различное направление. Следует различать следующие типы волокон: 1)волокна, проходящие в другое полушарие мозга через его спайки (мозолистое тело, передняя спайка, спайка свода) и направляющиеся к коре и базальным ядрам другой стороны (комиссуральные волокна); 2)системы волокон, соединяющих участки коры и подкорковые центры в пределах одной половины мозга (ассоциативные); 3) волокна, идущие от полушария большого мозга к нижележащим его отделам, к спинному мозгу и в обратном направлении от этих образований (проекционные волокна). Следующим отделом конечного мозга является мозолистое тело (corpus callosum), которое образовано комиссуральными волокнами, соединяющими оба полушария. Свободная верхняя поверхность мозолистого тела, обращенная в сторону продольной щели большого мозга, покрыта тонкой пластинкой серого вещества. Средняя часть мозолистого тела – его ствол – спереди загибается книзу, образуя колено мозолистого тела, которое, истончаясь, переходит в клюв, продолжающийся книзу в терминальную (пограничную) пластинку. Утолщенный задний отдел мозолистого тела заканчивается свободно в виде валика. Волокна мозолистого тела образуют в каждом полушарии большого мозга его лучистость. Волокна колена мозолистого тела соединяют кору лобных долей правого и левого полушарий. Волокна ствола соединяют серое вещество теменных и височных долей. В валике располагаются волокна, соединяющие кору затылочных долей. Участки лобной, теменной и затылочной долей каждого полушария отделены от мозолистого тела одноименной бороздой. Обратите внимание на то, что под мозолистым телом располагается тонкая белая пластинка – свод, состоящий из двух дугообразно изогнутых тяжей, соединенных в средней своей части поперечной спайкой свода (рис.). Тело свода, постепенно отдаляясь в передней части от мозолистого тела, дугообразно изгибается вперед и книзу и продолжается в столб свода. Нижняя часть каждого столба свода вначале подходит к терминальной пластинке, а далее столбы свода расходятся в латеральные стороны и направляются вниз и кзади, заканчиваясь в сосцевидных телах. Между ножками свода сзади и терминальной пластинкой спереди расположена поперечная передняя (белая) спайка, которая наряду с мозолистым телом соединяет между собой оба полушария большого мозга. Кзади тело свода продолжается в плоскую ножку свода, сращенную с нижней поверхностью мозолистого тела. Ножка свода постепенно уходит латерально и вниз, отделяется от мозолистого тела, еще больше уплотняется и одной своей стороной срастается с гиппокампом, образуя бахромку гиппокампа. Свободная сторона бахромки, обращенная в полость нижнего рога бокового желудочка, заканчивается в крючке, соединяя височную долю конечного мозга с промежуточным мозгом. Участок, ограниченный сверху и спереди мозолистым телом, снизу – его клювом, терминальной пластинкой и передней спайкой, сзади – ножкой свода, с каждой стороны занят сагиттально расположенной тонкой пластинкой – прозрачной перегородкой. Между пластинками прозрачной перегородки находится одноименная сагиттальная узкая полость, содержащая прозрачную жидкость. Пластинка прозрачной перегородки является медиальной стенкой переднего рога бокового желудочка. Рассмотрим строение внутренней капсулы (capsula internet)-толстой изогнутой под углом пластинки белого вещества, ограниченной с латеральной стороны чечевицеобразным ядром, а с медиальной – головкой хвостатого ядра (спереди) и таламусом (сзади). Внутренняя капсула образована проекционными волокнами, связующими кору большого мозга с другими отделами центральной нервной системы. Волокна восходящих проводящих путей, расходясь в различных направлениях к коре полушария, образуют лучистый венец. Книзу волокна нисходящих проводящих путей внутренней капсулы в виде компактных пучков направляются в ножку среднего мозга.
Рис.9. Свод мозга и гиппокамп. 1–мозолистое тело, 2–ядро свода, 3–ножка свода, 4–передняя спайка, 5–столб свода, 6–сосцевидное тело, 7–бахромка гиппокампа, 8–крючок, 9–зубчатая извилина, 10–парагиппокампальная извилина, 11–ножка гиппокампа, 12–гиппокамп, 13–боковой желудочек (вскрыт), 14–птичья шпора, 15–спайка свода.
Обратите внимание на то, что полостями полушарий большого мозга являются боковые желудочки (I и II), расположенные в толще белого вещества под мозолистым телом (рис. 11). У каждого желудочка выделяют четыре части: передний рог залегает в лобной доле, центральная часть – в теменной, задний рог – в затылочной, нижний рог – в височной доле. Передний рог обоих желудочков отделен от соседнего двумя пластинками прозрачной перегородки. Центральная часть бокового желудочка изгибается сверху вокруг таламуса, образует дугу и переходит кзади в задний рог, книзу – в нижний рог. Медиальной стенкой нижнего рога является гиппокамп (участок древней коры), соответствующий глубокой одноименной борозде на медиальной поверхности полушария. Медиально вдоль гиппокампа тянется бахромка, являющаяся продолжением ножки свода (рис.). На медиальной стенке заднего рога бокового желудочка мозга имеется выпячивание – птичья шпора, соответствующая шпорной борозде на медиальной поверхности полушария. В центральную часть и нижний рог бокового желудочка вдается сосудистое сплетение, которое через межжелудочковое отверстие соединяется с сосудистым сплетением III желудочка.
Рис.10. Проекция желудочков на поверхности большого мозга. 1–лобная доля, 2–центральная борозда, 3–боковой желудочек, 4–затылочная доля, 5–задний рог бокового желудочка, 6–IV желудочек, 7–водопровод мозга, 8–III желудочек, 9–центральная часть бокового желудочка, 10–нижний рог бокового желудочка, 11–передний рог бокового желудочка.
Рис.11. Фронтальный разрез головного мозга на уровне центральной части боковых желудочков. 1–центральная часть бокового желудочка, 2–сосудистое сплетение бокового желудочка, 3–передняя ворсинчатая артерия, 4–внутренняя мозговая вена, 5–свод, 6–мозолистое тело, 7–сосудистая основа III желудочка, 8–сосудистое сплетение III желудочка, 9–III желудочек, 10–таламус, 11–прикрепленная пластинка, 12–таламостриарная вена, 13–хвостатое ядро.
Кора больших полушарий Как указывалось выше, в состав конечного мозга человека входят два полушария, отделенных от друг друга продольной щелью. В глубине щели расположено соединяющее их мозолистое тело. Полушария головного мозга покрыты веществом, которое лежит по периферии (на поверхности) полушарий большого мозга. В коре головного мозга преобладает неокортекс (около 90%) – новая кора, которая возникла впервые у млекопитающих. Филогенетически более древние участки коры включают старую кору – архекортекс (зубчатая извилина и основание гиппокампа), а также древнюю кору – палеокортекс (препериформная, преамигдалярная и энторинальная области). Толщина коры в различных участках полушарий колеблется от 1,3 до 5 мм. Наиболее толстая кора находится в верхних участках предцентральной и постцентральной извилин и у парацентральной дольки. Кора выпуклой поверхности извилин толще, чем на боковых поверхностях и на дне борозд. Площадь поверхности коры больших полушарий взрослого человека достигает 450000 см2, 1/3 которой покрывает выпуклые части извилин и 2/3 – боковые и нижние стенки борозд. В коре содержится 10 – 14 млрд. нейронов, каждый из которых образует синапсы примерно с 8 – 10 тыс. других. Впервые отечественный ученый В.А.Бец показал, что строение и взаиморасположение нейронов не одинаково в различных участках коры, что определяет ее нейроцитоархитектонику. Клетки более или менее одинаковой структуры располагаются в виде отдельных слоев (пластинок). В новой коре большого мозга тела нейронов образуют шесть слоев. В различных отделах варьируют толщина слоев, характер их границ, размеры клеток, их количество и т.д. Высшим отделом ЦНС является кора большого мозга (кора больших полушарий). Она обеспечивает совершенную организацию поведения животных на основе врожденных и приобретенных в онтогенезе функций.
Морфофункциональная организация Кора большого мозга имеет следующие морфофункциональные особенности: - многослойность расположения нейронов; - модульный принцип организации; - соматотопическая локализация рецептирующих систем; - экранность, т. е. распределение внешней рецепции на плоскости нейронального поля коркового конца анализатора; - зависимость уровня активности от влияния подкорковых структур и ретикулярной формации; - наличие представительства всех функций нижележащих структур ЦНС; - цитоархитектоническое распределение на поля; - наличие в специфических проекционных сенсорных и моторной системах вторичных и третичных полей с ассоциативными функциями; - динамическая локализация функций выражается в возможности компенсаций функций утраченных структур; - перекрытие в коре большого мозга зон соседних периферических рецептивных полей; - возможность длительного сохранения следов раздражения; - реципрокная функциональная взаимосвязь возбудительных и тормозных состояний; - способность к иррадиации возбуждения и торможения; - наличие специфической электрической активности. Глубокие борозды делят каждое полушарие большого мозга на лобную, височную, теменную, затылочную доли и островок. Островок расположен в глубине сильвиевой борозды и закрыт сверху частями лобной и теменной долей мозга. Функции отдельных зон новой коры определяются особенностями ее структурно – функциональной организации, связями с другими структурами мозга, участием в восприятии, хранении и воспроизведении информации при организации и реализации поведения, регуляции функций сенсорных систем, внутренних органов. Особенности структурно – функциональной организации коры большого мозга обусловлены тем, что в эволюции происходила кортикализация функций, т.е. передача коре большого мозга функций нижележащих структур мозга. Однако эта передача не означает, что кора берет на себя выполнение функций других структур. Ее роль сводится к коррекции возможных нарушений функций взаимодействующих с ней систем, более совершенного с учетом индивидуального опыта анализа сигналов и организации оптимальной реакции на эти сигналы, формирование в своих и в других заинтересованных структурах мозга памятных следов о сигнале, его характеристиках, значении и характере реакции на него. В дальнейшем, по мере автоматизации реакция начинает выполняться подкорковыми структурами. В составе коры имеются пирамидные, звездчатые, веретенообразные нейроны. Пирамидные нейроны имеют разную величину, их дендриты несут большое количество шипиков, аксон пирамидного нейрона, как правило, идет через белое вещество в другие зоны коры или в структуры ЦНС. Звездчатые клетки имеют короткие хорошо ветвящиеся дендриты и короткий аксон, обеспечивающий связи нейронов в пределах самой коры большого мозга. Веретенообразные нейроны обеспечивают вертикальные или горизонтальные взаимосвязи нейронов разных слоев коры. Кора большого мозга имеет преимущественно шестислойное строение (рис.). Рассмотрим эти слои. Слой 1- верхний молекулярный, представлен в основном ветвлениями восходящих дендритов пирамидных нейронов, среди которых расположены редкие горизонтальные клетки и клетки – зерна, сюда же приходят волокна неспецифических ядер таламуса, регулирующие через дендриты этого слоя уровень возбудимости коры большого мозга. Слой 2 – наружный зернистый, состоит из звездчатых клеток, определяющих длительность циркулирования возбуждения в коре большого мозга, т. е. имеющих отношение к памяти. Слой 3 – наружный пирамидный, формируется из пирамидных клеток малой величины и вместе со 2 слоем обеспечивает корко- корковые связи различных извилин мозга. Слой 4 – внутренний зернистый, содержит преимущественно звездчатые клетки. Здесь заканчиваются специфические таламокорти-кальные пути, т. е. пути, начинающиеся от рецепторов анализаторов. Слой 5 – внутренний пирамидный, слой крупных пирамид, которые являются выходными нейронами, аксоны их идут в ствол мозга и спинной мозг. Слой 6 – слой полиморфных клеток, большинство нейронов этого слоя образуют кортико – таламические пути. Клеточный состав коры по разнообразию, морфологии, функции, формам связи не имеет себе равных в других отделах ЦНС. Нейронный состав, распределение нейронов по слоям в разных областях коры различны, что позволило выделить в мозге человека 53 цитоархитектонических поля. Разделение коры большого мозга на цитоархитектонические поля более четко формируется по мере совершенствования ее функции в филогенезе. У высших млекопитающих в отличие от низших от двигательного 4 поля хорошо дифференцируются вторичные поля 6,8 и 10, функционально обеспечивающие высокую координацию, точность движений; вокруг зрительного поля 17 – вторичные зрительные поля 18 и 19, участвующие в анализе значения зрительного стимула (организация зрительного внимания, управление движением глаза). Первичные слуховое, соматосенсорное, кожное и другие поля также имеют рядом расположенные вторичные и третичные поля, обеспечивающие ассоциацию функций данного анализатора с функциями других анализаторов. Для всех анализаторов характерен соматотопический принцип организации проекции на кору большого мозга периферических рецептирующих систем. Так, в сенсорной области коры второй центральной извилины имеются участки представительства, локализации каждой точки кожной поверхности, в двигательной области коры каждая мышца имеет свою топику (свое место), раздражая которую можно получить движение данной мышцы; в слуховой области коры имеется топическая локализация определенных тонов, повреждение локального участка слуховой области коры приводит к потере слуха на определенный тон. Особенностью корковых полей является экранный принцип их функционирования. Этот принцип заключается в том, что рецептор проецирует свой сигнал не на один нейрон коры, а на поле нейронов, которое образуется их коллатералями и связями. В результате сигнал фокусируется не точка в точку, а на множестве разнообразных нейронов, что обеспечивает его полный анализ и возможность передачи в другие заинтересованные структуры. Так одно волокно, приходящее в зрительную область коры, может активировать зону размером 0,1мм. Это значит, что один аксон распределяет свое действие на более чем 5000 нейронов. Входные (афферентные) импульсы поступают в кору снизу, поднимаются к звездчатым и пирамидным клеткам 3-5 слоев коры. От звездчатых клеток 4 слоя сигнал идет к пирамидным нейронам 3 слоя, а отсюда по ассоциативным волокнам – к другим полям, областям коры большого мозга. В коре входные и выходные элементы вместе со звездчатыми клетками образуют так называемые колонки - функциональные единицы коры, организованные в вертикальном направлении. Доказательством этого служит следующее: если микроэлектрод погружать перпендикулярно в кору, то на своем пути он встречает нейроны, реагирующие на один вид раздражения, если же микроэлектрод вводить горизонтально по коре, то он встречает нейроны, реагирующие на разные виды стимулов. Диаметр колонки около 500 мкм и определяется она зоной распределения коллатералей восходящего афферентного таламокортикального волокна. Соседние колонки имеют взаимосвязи, организующие участки множества колонок в организации той или иной реакции. Возбуждение одной из колонок приводит к торможению соседних. Каждая колонка имеет ряд ансамблей реализующих какую – либо функцию по вероятностно- статистическому принципу. Этот принцип заключается в том, что при повторном раздражении в реакции участвуют не вся группа нейронов, а её часть. Причем каждый раз часть участвующих нейронов может быть разной по составу, т. е. формируется группа активных нейронов (вероятностный принцип), среднестатистически достаточная для обеспечения нужной функции (статистический принцип). Как уже упоминалось, разные области коры большого мозга имеют разные поля, определяющиеся по характеру и количеству нейронов, толщине слоев и.т.д. Наличие структурно различных полей предполагает и разное их функциональное предназначение. Действительно, в коре большого мозга выделяют сенсорные, моторные и ассоциативные области. Электрические проявления активности коры большого мозга. Оценка функционального состояния коры большого мозга человека является трудной и до настоящего времени нерешенной задачей. Одним из признаков, косвенно свидетельствующем о функциональном состоянии структур головного мозга, является регистрация в них колебаний электрических потенциалов. Каждый нейрон имеет заряд мембраны, который при активации уменьшается, а при торможении – чаще увеличивается, т.е. развивается гиперполяризация. Глия мозга также имеет заряд мембран клеток. Динамика заряда мембраны нейронов, глии, процессы, происходящие в синапсах, дендритах, аксонном холмике, в аксоне – все это постоянно изменяющиеся, разнообразные по интенсивности, скорости процессы, интегральные характеристики которых зависят от функционального состояния нервной структуры и суммарно определяют ее электрические показатели. Если эти показатели регистрируются через микроэлектроды, то они отражают активность локального (100мкм в диаметре) участка мозга и называются фокальной активностью. В случае, если электрод располагается в подкорковой структуре, регистрируемая через него активность называется субкортикограммой, если электрод располагается в коре мозга - кортикограммой. Наконец, если электрод располагается на поверхности кожи головы, то регистрируется суммарная активность как коры, так и подкорковых структур. Это проявление активности называется электроэнцефалограммой (ЭЭГ). Все виды активности мозга в динамике подвержены усилению и ослаблению и сопровождаются определенными ритмами электрических колебаний. У человека в покое при отсутствии внешних раздражений преобладают медленные ритмы изменения состояния коры мозга, что на ЭЭГ находит отражение в форме так называемого альфа-ритма, частота колебаний которого составляет 8-13 в секунду, а амплитуда – приблизительно 50 мкВ. Переход человека к активной деятельности приводит к смене альфа-ритма на более быстрый бета- ритм, имеющий частоту колебаний 14-30 в секунду, амплитуда которых составляет 25 мкВ. Переход от состояния покоя к состоянию сосредоточенного внимания или ко сну сопровождается развитием боле медленного тета – ритма (4-8 колебаний в секунду) или дельта – ритма (0,5- 3,5 колебаний в секунду). Амплитуда медленных ритмов составляет 100-300 мкВ. Когда на фоне покоя или другого состояния мозгу предъявляется новое быстрое нарастающее раздражение, на ЭЭГ регистрируются так называемые вызванные потенциалы (ВП). Они представляют собой синхронную реакцию множества нейронов данной зоны коры. Латентный период, амплитуда ВП зависят от интенсивности наносимого раздражения. Компоненты ВП, количество и характер его колебаний зависят от адекватности стимула относительно зоны регистрации ВП. ВП может состоять из первичного ответа или же из первичного и вторичного. Первичные ответы представляют собой двухфазные, позитивно- негативные колебания. Они регистрируются в первичных зонах коры анализатора и только при адекватном для данного анализатора стимуле. Например, зрительная стимуляция для первичной зрительной коры (поле 17) является адекватной. Первичные ответы характеризуются коротким латентным периодом (ЛП), двухфазностью колебаний: вначале положительная, затем отрицательная фазы. Первичный ответ формируется за счет кратковременной синхронизации активности близлежащих нейронов. Вторичные ответы более вариабельны по ЛП, длительности, амплитуде, чем первичные. Как правило, вторичные ответы чаще возникают на сигналы, имеющие определенную смысловую нагрузку, на адекватные для данного анализатора стимулы; они хорошо формируются при обучении.
|
||||||||||||
|
|
Последнее изменение этой страницы: 2017-01-20; просмотров: 1160; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.12.165.68 (0.017 с.) |


 Скорлупа
Скорлупа Бледный шар
Бледный шар Нижняя поверхность переднего отдела чечевицеобразного ядра прилежит к переднему продырявленному веществу и соединяется с хвостатым ядром. Медиальная часть чечевицеобразного ядра на горизонтальном разрезе головного мозга суживается и углом обращена к колену внутренней капсулы, находящемуся на границе таламуса и головки хвостатого ядра. Выпуклая латеральная поверхность чечевицеобразного ядра обращена к основанию островковой доли полушария большого мозга.
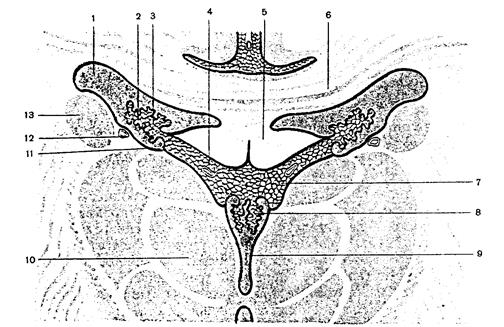
Нижняя поверхность переднего отдела чечевицеобразного ядра прилежит к переднему продырявленному веществу и соединяется с хвостатым ядром. Медиальная часть чечевицеобразного ядра на горизонтальном разрезе головного мозга суживается и углом обращена к колену внутренней капсулы, находящемуся на границе таламуса и головки хвостатого ядра. Выпуклая латеральная поверхность чечевицеобразного ядра обращена к основанию островковой доли полушария большого мозга. 1–кора большого мозга (плащ), 2–колено мозолистого тела, 3–передний рог бокового желудочка, 4–внутренняя капсула, 5–наружная капсула, 6–ограда, 7–самая наружная капсула, 8–скорлупа, 9–бледный шар, 10–III желудочек, 11–задний рог бокового желудочка, 12–зрительный бугор, 13–корковое вещество (кора) островка, 14–головка
1–кора большого мозга (плащ), 2–колено мозолистого тела, 3–передний рог бокового желудочка, 4–внутренняя капсула, 5–наружная капсула, 6–ограда, 7–самая наружная капсула, 8–скорлупа, 9–бледный шар, 10–III желудочек, 11–задний рог бокового желудочка, 12–зрительный бугор, 13–корковое вещество (кора) островка, 14–головка