Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Лекция 6 от 30 сентября 2008 г.Содержание книги
Поиск на нашем сайте
Лекция 6 от 30 сентября 2008 г. Межклеточная передача возбуждения [Мф1] . Общая физиология синапсов.
Мы продолжаем рассматривать вопросы распространения возбуждения. Но если на предыдущей лекции мы ограничились проблемой передачи возбуждения между отдельными частями клетки, причём основной темой являлось распространение возбуждения вдоль волокна, то сегодня мы разберём вопросы передачи возбуждения от одной клетки к другой. Ещё в 60-х годах прошлого столетия И.М.Сеченов подчеркивал, что вне рассмотрения межклеточной связи нельзя объяснить способы происхождения даже самого элементарного нервного процесса, т.е. информационного процесса[Мф2]. В центре нашего внимания будет синапс.
План лекции
1. Понятия «синапс», «нексус». 3 2. Классификация синапсов. 3 По характеру контактирующих клеток. 3 По способу передачи сигнала. 3 По конечному эффекту. 3 По форме контакта. 4 По развитию в онтогенезе. 4 Типы межнейрональных синапсов. 4 По характеру контактирующих частей нейронов 4 По локализации в нервной системе. 4 3. Химический синапс: общий план строения 5 4. Типы химических синапсов. 6 5. Этапы передачи сигнала в химическом синапсе 6 Везикулярная гипотеза освобождения медиатора в синапсе 6 Формирование везикул. 7 Заполнение везикул. 7 Опустошение везикул и освобождение медиатора 8 Рециклизация везикул. 9 Кальциевые каналы и экзоцитоз. 9 6. Медиаторы химического синапса. 9 Понятие «медиатор химического синапса». 9 Доказательство возможности химической передачи возбуждения. Опыт О.Лёви. 9 Виды (классификация) медиаторов химического синапса 10 Принцип Г.Х.Дейла. 10 Происхождение медиаторов химического синапса 11 7. Комедиаторы, модуляторы, агонисты, антагонисты. 11 Комедиаторы (сомедиаторы) 11 Модуляторы.. 12 Агонисты.. 12 Антагонисты.. 12 8. Ионотропные рецепторы.. 12 Постсинаптические потенциалы.. 13 9. Метаботропные синапсы.. 14 Структура и функция G-белка при передаче сигнала 15 ГТФ-азный цикла G‑ белков. 15 10. Нервно-мышечный синапс скелетного мышечного волокна 18 Структура нервно-мышечного синапса. 19 Рецепторы постсинаптической мембраны.. 20 Варианты изображения Н-холинорецептора (nicotinic acetylcholine receptors) 21 Этапы передачи возбуждения в НМС.. 22
Миниатюрный потенциал концевой пластинки 22 Фармакологическая модификации НМС.. 22 11. Электрические синапсы. Смешанные синапсы. 22 Сравнительная характеристика химического и электрического синапсов 23 Рекомендуемая литература. 24 Основная. 24 Дополнительная. 24 Приложение 1. Межнейрональные синапсы. 24 Приложение 2. Биохимия медиаторов. 24 Приложение 3. Холинергические синапсы.. 25 Приложение 4. Адренергические синапсы.. 25 Приложение 5. Термины.. 25 Методические указания. 26
Понятия «синапс», «нексус»
Дадим определение нексусам.
Классификация синапсов
Синапсы можно разделить, положив в основу различные критерии.
Иногда в литературе встречается термин «внутриклеточный синапс». [3] Например, так называют место, где Т‑трубочка близко подходит к терминальной цистерне саркоплазматического ретикулюма мышечного волокна. [A12]
Соединение между аксонами нейросекреторных клеток гипоталямуса и стенкой капилляра, обеспечивающее поступление нейрогормона в кровь, тоже называют синапсом, вазоневральным, или нейрокапиллярным, или аксоваскулярным. Однако под классическое понятие «синапс» этот тип соединения, безусловно, не подпадает.
На рис. 410021125 проходящие синапсы изображены более детально.
Типы химических синапсов
Возбуждающими и тормозными могут быть и электрические синапсы. Но у человека тормозных электрических синапсов не найдено, а функция возбуждающих и тормозных химических синапсов у человека хорошо изучена.
5. Этапы передачи сигнала в химическом синапсе[Мф24]
При огромном разнообразии синапсов и различиях в механизмах передачи сигнала в них, для большинства изученных синапсов можно выделить следующие наиболее общие этапы передачи возбуждения:
Везикулярная гипотеза освобождения медиатора в синапсе [A25]
Для того чтобы понять, как формировалась везикулярная гипотеза, необходимо остановиться на квантовой гипотезе освобождения медиатора в синапсе.
В 1953 году Бернард Катц с сотрудниками, исследуя нервно-мышечный синапс лягушки (медиатор - ацетилхолин), с помощью микроэлектродов зарегистрировали новый класс постсинаптических потенциалов (рис. 809260319).
Эти сигналы возникали случайно, в покое, имели очень маленькую амплитуду и были названы миниатюрными потенциалами концевой пластинки (МПКП). Далее оказалось, что постсинаптические потенциалы, вызванные раздражением двигательного нерва (потенциалы концевой пластинки - ПКП), от раздражения к раздражению варьируют по амплитуде, причем эти колебания кратны амплитуде МПКП. Было предположено, что медиатор в синапсе освобождается в виде мультимолекулярных порций - квантов. В покое случайное освобождение из нервного окончания отдельных порций вызывает появление на постсинаптической мембране МПКП, а в ответ на раздражение происходит синхронное освобождение нескольких десятков или сот квантов и возникает ПКП. Электрофизиологическое определение показало, что квант медиатора состоит из 1000-10 000 молекул ацетилхолина. В дальнейшем квантовая гипотеза освобождения медиатора получила подтверждение на самых разнообразных объектах и в настоящее время считается общепризнанной.
Использование метода электронной микроскопии для изучения ультраструктуры синапса позволило в 1954 году Де Робертсу и Беннету выявить в цитоплазме двигательного нервного окончания большое количество синаптических везикул диаметром около 50 нм. Поскольку везикулы имели одинаковые размеры и концентрировались у пресинаптической мембраны, было предположено, что квант медиатора находится в синаптической везикуле, а освобождение медиатора происходит путем выделения содержимого везикулы в синаптическую щель путем экзоцитоза. Так была сформулирована везикулярная гипотеза освобождения медиатора в синапсе. В дальнейшем синаптические везикулы были обнаружены во всех химических синапсах нервной системы. К основным постулатам везикулярной гипотезы необходимо отнести следующие: 1) медиатор в нервном окончании концентрируется в синаптических везикулах, 2) везикулярный медиатор освобождается путем слияния мембраны везикулы с пресинаптической мембраной (экзоцитоз).
Типы везикул. Как показали электронно-микроскопические исследования, самые разнообразные нервные окончания имеют два типа секреторных везикул: синаптические везикулы (мелкие везикулы) и секреторные гранулы (так называемые крупные или электронно-плотные везикулы). Мелкие синаптические везикулы однородны по размерам и имеют малый диаметр (около 50 нм). Эти везикулы содержат классические медиаторы. Крупные, электронно-плотные везикулы имеют большой диаметр (около 100 нм), они неоднородны по размерам и содержат электронно-плотные гранулы, представляющие собой крупномолекулярные медиаторы - пептиды и белки.
Формирование везикул Синаптические везикулы образуются в теле нервной клетки из эндоплазматического ретикулума и цистерн аппарата Гольджи, а затем транспортируются по аксону в нервные окончания.
По всей видимости, принципиальной разницы в формировании различных типов везикул, заполненных различными медиаторами, не наблюдается.
Заполнение везикул
Как показали биохимические исследования, медиатор в мелких синаптических везикулах находится в очень высокой концентрации - 100 ммоль/л. Это достигается наличием в мембране везикулы специальных активных транспортных систем. В мембране везикулы имеется протонный насос, который, используя энергию АТФ, создает разность потенциалов на мембране везикулы (содержимое везикулы заряжено положительно по сравнению с цитоплазмой нервного окончания). Везикулы содержат также хлорные каналы. Электрохимический градиент, формируемый протонным насосом, обеспечивает активный транспорт медиатора, который синтезируется в цитоплазме нервного окончания, в везикулу. В настоящее время выделены несколько классов таких транспортных молекул, специфичных для биогенных аминов, ацетилхолина, глютамата, ГАМК и глицина. Наряду с медиатором в везикулах находятся АТФ, ионы, ферменты и другие компоненты.
Что касается крупных, электронно-плотных везикул, то их заполнение белковыми компонентами начинается уже в процессе образования везикул из эндоплазматического ретикулума. Синтез нейроактивных пептидов происходит подобно синтезу пептидных гормонов. Первоначально крупные аминокислотные последовательности (пептидные цепи) образуются на рибосомах, подобно прегормонам, и помещаются в эндоплазматический ретикулум. В цистернах аппарата Гольджи начинается протеолитический процесс разделения крупных полипептидов на фрагменты с образования активных пептидов, которые включаются в отпочковывающиеся везикулы. Причем разные нейроактивные пептидные фрагменты могут оказаться в различных везикулах, которые транспортируются в нервные окончания нейрона. Во многих нейронах медиаторы и нейропептиды синтезируются и упаковываются в одни и те же везикулы, следовательно, из их нервных окончаний освобождаются несколько различных медиаторов.
Рециклизация везикул
Под процессом рециклизации понимают процесс повторного образования, заполнения и использования везикул после их опустошения и экзоцитоза. Если считать, что освобождение медиатора происходит путем экзоцитоза с полным слиянием, то естественно, что при активности синапса будет происходить уменьшение количества и в конечном итоге исчезновение везикул. Однако в нервном окончании наряду с процессом экзоцитоза осуществляется также эндоцитоз, то есть процесс образования новых везикул из фрагментов пресинаптической мембраны. Для мелких везикул этот процесс заключается в том, что после экзоцитоза из фрагмента пресинаптической мембраны путем эндоцитоза образуется везикула, которая через стадию эндосомы вновь заполняется медиатором и может участвовать в освобождении медиатора. В модели экзоцитоза без полного слияния рециклизация заключается в отхождении опустошенной везикулы от пресинаптической мембраны и повторном заполнении ее медиатором. Процесс рециклизации везикул достаточно быстрый и занимает не более 1 мин. Крупные, электронно-плотные везикулы, заполненные пептидами, не могут восстанавливаться после экзоцитоза.
Рис. 30110005161. Схема опыта О.Лёви (Loewi). При раздражении n.vagus сердца 1 выделялся медиатор, действовавший на сердце 2.
|


 Английский нейрофизиолог Чарльз С.Шеррингтон (Sir Charles Scott Sherrington) (1857[Б6] -1952) в 1932 г. был удостоен Нобелевской премии[Б7]. Школа Шеррингтона заложила основы современной нейрофизиологии (Гранит Р., Лиддел Дж.Т., Магнус Р., Пенфилд У., Экклс Дж.).
Английский нейрофизиолог Чарльз С.Шеррингтон (Sir Charles Scott Sherrington) (1857[Б6] -1952) в 1932 г. был удостоен Нобелевской премии[Б7]. Школа Шеррингтона заложила основы современной нейрофизиологии (Гранит Р., Лиддел Дж.Т., Магнус Р., Пенфилд У., Экклс Дж.).

 Рис. 410020742. А — терминальные синапсы (ТС); Б — проходящие синапсы (ПС).
Рис. 410020742. А — терминальные синапсы (ТС); Б — проходящие синапсы (ПС).
 Рис. 410021125. Проходящие синапсы[Мф17].
Рис. 410021125. Проходящие синапсы[Мф17].
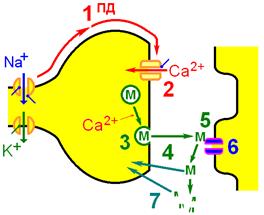



 Рис. 809300818. Формирование, заполнение и опустошение везикул (а – мелких, б – крупных).
Рис. 809300818. Формирование, заполнение и опустошение везикул (а – мелких, б – крупных).
 Рис. 809300905. Рециклизация везикул
Рис. 809300905. Рециклизация везикул
Лёви О. принадлежит открытие фермента, который гидролизует ацетилхолин, позднее названный холинэстеразой.
Медиаторную функцию выполняет большая группа разнородных химических веществ. Известно более 50 химических веществ[Мф28]. Список вновь открываемых химических медиаторов неуклонно пополняется[Б29].
Чаще всего химическими медиаторами являются вещества с небольшой относительной молекулярной массой. Однако и высокомолекулярные соединения, такие, как полипептиды, также способны выполнять роль химических передатчиков в ряде центральных и периферических синапсов.
Основные критерии медиаторной функции веществ: 1. наличие вещества в соответствующих пресинаптических окончаниях; 2. способность вещества высвобождаться под влиянием потенциала действия; 3. идентичность молекулярных и ионных механизмов действия на постсинаптическую мембрану вещества, высвобождаемого потенциалом действия и прикладываемого искусственно к постсинаптической мембране;
|
В периферических структурах процесс идентификации медиатора по указанным выше критериям может быть произведен сравнительно просто. ЦНС построена из негомогенных диффузно расположенных популяций нервных клеток и окончаний. Это вносит значительные трудности в обнаружение выделяемого медиатора, который, прежде чем появиться на поверхности мозга или в спинномозговой жидкости, должен диффундировать на большие расстояния. Помимо этого, в центральных структурах трудно добиться избирательной стимуляции определенной гомогенной группы нейронов или волокон, так же как трудно подводить предполагаемый медиатор к определенным нервным клеткам, не оказывая влияния на соседние нейроны. Именно поэтому природа химических медиаторов в периферических структурах нервной системы достаточно хорошо изучена, а в ЦНС до сих пор окончательно не установлена. Тем не менее, выявлен ряд веществ, играющих роль медиаторов синаптического возбуждения и торможения в ЦНС млекопитающих и человека.
Принцип Г.Х.Дейла
Согласно принципу Дейла, каждый нейрон во всех своих синаптических окончаниях выделяет один и тот же медиатор[Б33]. Или другими словами – «один нейрон – один медиатор». Иллюстрация принципа Дейла: периферические окончания аксонов моторных нейронов активируют волокна скелетной мускулатуры с помощью ацетилхолина. Возвратные коллатерали тех же аксонов в пределах ЦНС выделяют тот же медиатор.
Рис. 8092308321. Принцип Дейла. Объяснения в тексте.
|

ДЕЙЛ (Dale) Генри Халлетт (1875-1968), английский физиолог и фармаколог, иностранный почетный член АН СССР (1942). Исследовал химическую природу передачи нервного импульса через синапсы и роль в ней ацетилхолина. Нобелевская премия «за открытие химической природы передачи нервной реакции» (1936, совместно с О.Лёви[Б34]).
Однако, в настоящее время считают, что принцип Дейла имеет исторический интерес или по меньшей мере имеется большое количество исключений из этого принципа.
Что противоречит принципу Дейла?
Так в вегетативной нервной системе, по крайней мере у эмбрионов, одни и те же нейроны высвобождают как ацетилхолин, так и адреналин. В двигательной концевой пластинке вместе с ацетилхолином выделяется аденозинтрифосфат, который, вероятно, также служит медиатором. Часто из синаптического окончания высвобождаются одновременно классический медиатор, например норадреналин, и участвующий в нервной передаче пептид. Особенности такого совместного действия медиаторов (сомедиаторов) пока неясны, но его эффект, вероятно, чаще всего сводится к определенному типу модуляции.
Тем не менее, принятие физиологами принципа Дейла способствовало появлению традиции обозначать нейроны по типу медиатора, который выделяют их окончания. Нейроны, освобождающие ацетилхолин, называют холинергическими, серотонин — серотонинергическими и т.д. Этот же принцип может быть использован для обозначения различных химических синапсов. Иными словами, различают холинергические, серотонинергические и другие синапсы. Теперь мы знаем, что придерживаться этой традиции надо очень осторожно.
Следует также признать, что в синапсах, как правило, можно выделить доминирующий медиатор и, следовательно, говорить об относительной специфичности синапса[Б35].
Рис. 8092308321. Происхождение медиаторов химического синапса. Объяснения в тексте.
|
После каждого цикла проведения импульса медиатор разрушается, либо идет обратный захват в пресинаптическую структуру. В одних случаях захватывается неразрушенный медиатор, в других - его осколки.
Ионотропные рецепторы

| [18] |
Ионотропные рецепторы содержат домен, узнающий медиатор и ионный канал. Функциональная структура хемовозбудимого канала схематически показана на рис. 709192337, 809250525.
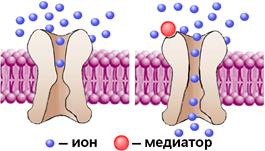 Рис. 709192337. Схематическое изображение хемовозбудимого ионного канала. До взаимодействия молекулы медиатора с рецептором ворота закрыты (слева). Они открываются при связывании медиатора рецептором (справа).
Рис. 709192337. Схематическое изображение хемовозбудимого ионного канала. До взаимодействия молекулы медиатора с рецептором ворота закрыты (слева). Они открываются при связывании медиатора рецептором (справа).
|
 Рис. 809250525. Упрощённое схематическое изображение хемовозбудимого ионного канала (смотри рис. 709192337).
Рис. 809250525. Упрощённое схематическое изображение хемовозбудимого ионного канала (смотри рис. 709192337).

|

Хемовозбудимые натриевые каналы постсинаптических мембран имеют только один воротный механизм, активируемый медиатором.
Вспомните! Потенциалзависимые натривые каналы, обеспечивающие развитие регенеративной деполяризации (спайка потенциала действия) имеют два воротных механизма – активационный и инактивационный.
В отсутствие медиатора канал закрыт. Взаимодействие медиатора с рецептором приводит к активации натриевого канала. В результате мембрана деполяризуется.
В качестве примера ионотропного синапса приведем Н-холинергический синапс (рис. 809250501).
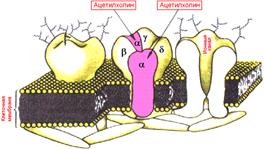 Рис. 809250501. Локализация действия ацетилхолина на модели Н‑холинорецептора.
Рис. 809250501. Локализация действия ацетилхолина на модели Н‑холинорецептора.
|
Рецепторы постсинаптических мембран - это белковые структуры, которые являются интегральными белками плазматической мембраны, они синтезируются в эндоплазматическом ретикулюме клетки, после чего они попадают (встраиваются) в постсинаптическую мембрану, пройдя предварительно "сортировку" в аппарате Гольджи.При нарушении белкового синтеза концентрация рецепторов может существенно снижаться и приводить к уменьшению функциональных возможностей синаптической передачи. В ряде случаев у человека могут вырабатываться антитела к собственным рецепторам постсинаптической мембраны. Это приводит к нарушению функции синапса. Синтез рецепторов - контролируемый процесс. Например, если мышца денервирована, то число рецепторов области постсинаптической мембраны снижается. Одновременно во внесинаптических областях возрастает концентрация рецепторов, это делает мышцу более чувствительной в отношении циркулирующего в крови медиатора [Б38].
Метаботропные синапсы
|
| [19] |
При передаче сигнала в метаботропных синапсах используется мультимолекулярная система (рис. 709210836), состоящая из
· рецептора,
· ГТФ-связывающего белка (G-белка),
· фермента,
· второго посредника,
· эффектора.
 Рис. 709210836. Мультимолекулярная система: рецептор – G-белок - фермент - второй посредник - … - эффектор. 1 ‑ взаимодействие медиатора и рецептора; 2 ‑ активация рецептором G-белка; 3 ‑ активация альфа-субединицей фермента; 4 ‑ образование второго посредника; 5 ‑ влияние на ионный канал; 6 ‑ влияние на транскрипцию или трансляцию.
Рис. 709210836. Мультимолекулярная система: рецептор – G-белок - фермент - второй посредник - … - эффектор. 1 ‑ взаимодействие медиатора и рецептора; 2 ‑ активация рецептором G-белка; 3 ‑ активация альфа-субединицей фермента; 4 ‑ образование второго посредника; 5 ‑ влияние на ионный канал; 6 ‑ влияние на транскрипцию или трансляцию.
|
 Рис. 809250915. Мультимолекулярная система: рецептор – G-белок - фермент - второй посредник - … - эффектор (смотри рис. 709210836, обозначения те же).
Рис. 809250915. Мультимолекулярная система: рецептор – G-белок - фермент - второй посредник - … - эффектор (смотри рис. 709210836, обозначения те же).
|
| Как могут быть связаны (сопряжены) метаботропные рецепторы с ионным каналом? 1. Через G-белки (система «G-белок – ионный канал»): М‑холинорецепторы, адренорецепторы 2. Через изменение концентрации внутриклеточного посредника типа циклического аденозинмонофосфата (цАМФ), в результате чего меняется активность внутриклеточных протеаз - а как следствие этого - изменение функциональной активности клетки. |
Рис. 709200023. Модуляция потенциал-зависимых кальциевых каналов норадреналином.
[Мф45] Пример регуляции экспрессии белков путем активации метаботропных рецепторов показан на рис. 809251517.
Рис. 809251517. Регуляция экспрессии белков путем активации метаботропных рецепторов.
|
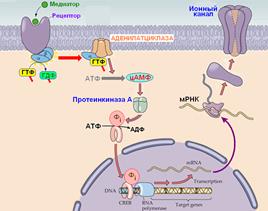
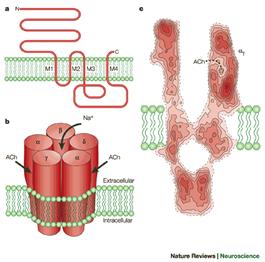
Рис. 809252331. Никотиновый рецептор, формирующий ионный канал.
Две a-субъединицы содержат два связывающих ацетилхолин сайта, ассоциированных с каналом. При взаимодействии этих сайтов с ацетилхолином значительно увеличивается вероятность открытия канала. Три других субъединицы b-субъединица, g-субъединица и d-субъединица доукомплектовывают протеин. Субъединицы, полипептидные цепи которых четыре раза пронизывают липидный бислой, с внешней стороны гликозилированы, а внутри взаимодействуют с белками тубулинового и актинового цитоскелета.
Электронная микроскопия показывает, что никотиновый рецепторный канал - это симметричная структура, похожая на цилиндр, живой поясок с каналом в центре. Приблизительно по одной трети протеина выступает с обеих сторон плазмалеммы наружу и внутрь клетки, остальная часть находится в толще мембраны. Наружный выступ напоминает воронку, являющуюся входом в канал.
Никотиновые каналы обладают лишь относительной специфичностью. Они обеспечивают диффузию любых катионов. Поскольку покоящаяся мембрана имеет потенциал близкий к равновесному потенциалу для K+, то это означает, что будет диффундировать много больше ионов натрия Na+ и кальция Ca2+ внутрь клетки, чем калия K+ наружу. Это является причиной деполяризации постсинаптической мембраны и возбуждения мышечной клетки. В отличие от других синапсов, этот вид деполяризации получил название потенциала концевой пластинки (ПКП).
Варианты изображения
Н-холинорецептора
(nicotinic acetylcholine receptors)
|
| [33] |
 http://www.nature.com/nrn/journal
http://www.nature.com/nrn/journal
|
 http://journals.cambridge.org/
http://journals.cambridge.org/
|
 http://media-2.web.britannica.com/eb-media
http://media-2.web.britannica.com/eb-media
|
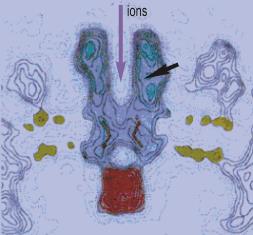 Рецептор изображен синим, фосфолипидные головки мембраны желтым и прикрепленный цитоплазматический белок розовым. Рецептор приблизительно 12 нм длиной выдвинут приблизительно на 6.5 нм во внеклеточное и на 1.5 нм во внутриклеточное пространство. Узкая пора (длинная стрелка) диаметром ~2 нм сформирована кольцом из пяти aльфа-спиралей. Участок связывания АХ выглядит как карман (короткая стрелка) в aльфа-субъединице и расположен приблизительно в 5 нм от ворот. (По данным Найджела Унвина, MRC Лаборатория Молекулярной Биологии, Кембриджа.)
http://medbiol.ru/medbiol/cell_sign3/00033e4a.htm
Рецептор изображен синим, фосфолипидные головки мембраны желтым и прикрепленный цитоплазматический белок розовым. Рецептор приблизительно 12 нм длиной выдвинут приблизительно на 6.5 нм во внеклеточное и на 1.5 нм во внутриклеточное пространство. Узкая пора (длинная стрелка) диаметром ~2 нм сформирована кольцом из пяти aльфа-спиралей. Участок связывания АХ выглядит как карман (короткая стрелка) в aльфа-субъединице и расположен приблизительно в 5 нм от ворот. (По данным Найджела Унвина, MRC Лаборатория Молекулярной Биологии, Кембриджа.)
http://medbiol.ru/medbiol/cell_sign3/00033e4a.htm
|
Рис. 070306121. Структура электрического синапса. Объяснение в тексте
Рис.. Схема чисто электрического синапса между дендритами мотонейронов лягушки (A), смешанного синапса между центральными окончаниями первичного афферентного нейрона и мотонейроном лягушки (B) и химического синапса между центральными окончаниями первичного афферентного нейрона и мотонейрона кошки (C). Основная
Физиология человека / Под редакцией В.М.Покровского, Г.Ф.Коротько Медицина, 2003 (2007) г. С. 69-74.
Физиология человека: Учебник / В двух томах. Т.I / В.М.Покровский, Г.Ф.Коротько, В.И.Кобрин и др.; Под ред. В.М.Покровского, Г.Ф.Коротько.— М.: Медицина, 1998.— [Б54] С.66-71, 90-91.
Дополнительная 1. Основы физиологии человека. В 2-х т. Т.I / Под ред. Б.И.Ткаченко. - СПб, 1994. - [Б55] С.101-108, 118-124.. 2. Общий курс физиологии человека и животных. В 2 кн. Кн.1. Физиология нервной, мышечной и сенсорной систем: Учебн. для биол. и мед. спец. вузов / А.Д.Ноздрачев, И.А.Баранникова, А.С.Батуев и др.; Под ред. А.Д.Ноздрачева.- М.: Высш. шк. 1991.- С.56-68[Б56]. 3. Физиология человека / Под ред. Г.И.Косицкого.- 3-е изд., перераб. и доп.- М.: Медицина, 1985.- С.92-102, 74-81,[Б57] 82-84. 4. Физиология человека: В 3-х томах. Т.1. Общая физиология клетки. Интегративная функция нервной системы. Физиология мышц. Сенсорная физиология. Пер. с англ. / Под ред. Р.Шмидта и Г.Тевса.- Изд. 2-е, доп. и перераб.- М.: Мир, 1996.- C. 51-68 С.[Б58]. 5. Котляр Б.И., Шульговский В.В. Физиология центральной нервной системы.- М.: Изд-во Моск. ун-та, 1979.- С.55-77[Б59]. 6. Кузнецов Атлас по гистологии С.111-113
Приложение 1. Межнейрональные синапсы.
Рис.. Аксодентритический синапс
Рис.. Аксоаксональный (ААС) и аксосоматический (АСС) синапсы.
Рис.. Аксосоматические (АСС) синапсы. АЗ — активная зона.
Приложение 2. Биохимия медиаторов
Ацетилхолин. Ацетилхолин является уксуснокислым эфиром холина, т.е. относится к простым эфирам. Он образуется при ацетилировании холина, причем этот процесс происходит при участии фермента ацетилхолинтрансферазы. Особенностью ацетилхолина как медиатора является быстрое его разрушение после высвобождения из пресинаптических окончаний с помощью фермента ацетилхолинэстеразы. Катехоламины. Три родственных в химическом отношении вещества: дофамин, норадреналин и адреналин — являются производными тирозина. Последовательно образуются ДОФА, дофамин, норадреналин - адреналин). Серотонин. Подобно катехоламинам, серотонин относится к группе моноаминов, синтезируется из аминокислоты триптофана. Нейтральные аминокислоты. Две основные дикарбоксильные кислоты L-глутамат и L-аспартат. L-глутаминовая кислота представляет собой дикарбоновую аминокислоту, входящую в состав многих белков и пептидов. Она плохо проходит через гематоэнцефалический барьер и поэтому не поступает в мозг из крови, образуясь в самой нервной ткани (главным образом из глюкозы). Кислые аминокислоты. К этой группе аминокислот относятся ГАМК и глицин. ГАМК представляет собой продукт декарбоксилирования L-глутаминовой кислоты. Эта реакция катализируется декарбоксилазой глутаминовой кислоты. Отмечено значительное совпадение локализации этого фермента и ГАМК в пределах ЦНС. Другой фермент нервной ткани — трансаминаза — катализирует перенос аминогруппы ГАМК на a-кетоглутаровую кислоту, в результате чего последняя превращается в семиальдегид янтарной кислоты. Под названием «вещество Р» подразумевается группа агентов, впервые экстрагированных из кишечника.
Приложение 3. Холинергические синапсы
Этот тип синапсов, особенно мионевральный хорошо изучены.
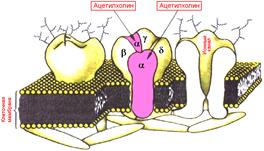
Рис. 3.3. Локализация действия ацетилхолина на модели н-холинорецептора.
Фармакологически было показано, что в одних структурах холинорецепторы (ХР) активируются помимо ацетилхолина ( АХ), также и никотином, другие же рецепторы, помимо ацетилхолина способные активироваться мускарином (алкалоид ряда грибов, в том числе мухомора). В связи с этим все холинорецепторы разделены на 2 основных кла
|
||||
|
|
Последнее изменение этой страницы: 2017-01-19; просмотров: 693; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.148.115.187 (0.018 с.) |