Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Лекция №1. Введение. Предмет молекулярной биологии. История развития. Эволюция клетки. Биологическое значение нуклеиновых кислот.Содержание книги
Поиск на нашем сайте
РАЗДЕЛ 1. ГЛОССАРИЙ Автотрофы – организмы, синтезирующие нужные им органические вещества из неорганических соединений. Автогамия — самооплодотворение как форма полового размножения у простейших путем слияния гаплоидных ядер в одной зародышевой клетке. Адаптация — приспособление. Аденин – пурин, входящий в состав нуклеиновых кислот и нуклеотидов, играющих важную роль в переносе энергии. Аденозинтрифосфат (АТФ) – органическое вещество, содержащее аденин, рибозу и три фосфатные группы; играет важнейшую роль в переносе энергии в биологических системах. Активный перенос – перенос веществ в клетку или из клетки через плазматическую мембрану против градиента концентрации с помощью механизма, требующего затраты энергии. Аллель – один из нескольких альтернативных вариантовгена, которые могут находиться в данном участке (локусе) хромосомы. Аминокислота – органическое соединение, содержащее аминогруппу и карбоксильную группу; аминокислоты могут соединяться между собой, образуя пептидные цепи белковых молекул. Анаболизм – биохимические реакции, в результате которых из более простых веществ синтезируются более сложные, что приводит к запасанию энергии, к образованию новых материалов для построения клеток и к росту. Аналогичные (органы) — сходные по функции, но разные по происхождению. Анафаза – стадия митоза или мейоза, на которой хромосомы расходятся к полюсам веретена; следует за метафазой. Анаэробный — безвоздушный; термин относится к организмам, существующим в бескислородной среде. Антиген – чужеродное вещество, обычно белок или комплекс полисахарида с белком, которое, попадая в организм, вызывает образование специфических антител. Антитело – специфический белок, вырабатываемые организмом в ответ на появление в крови или тканях чужеродного вещества. Апомиксис — размножение без оплодотворения при партеногенезе. Апоморфный — признак отражает эволюционно продвинутое состояние морфологии органов. Ароморфоз — тип биологического прогресса в эволюции, который приводит к повышению морфофизиологической организации организмов. Аутосомы – обычные парные хромосомы в отличие от половых хромосом. Ацеломический — не имеющий вторичной полости тела — целома. Бактерии – очень мелкие, обычно одноклеточные, микроорганизмы, для которых характерно отсутствие оформленного ядра. Белки – макромолекулярные вещества, содержащие углерод, водород. Кислород, азот и обычно также серу и фосфор. Построены из аминокислот, соединенных в цепи пептидными связями. Бентос — организмы, обитающие на дне водоемов. Бесполое размножение — форма размножения, не включающая мейоз и слияние гамет. Билатеральная симметрия — тип симметрии, при котором через тело животного можно провести лишь одну плоскость симметрии, делящую его на две идентичные половины. Биогеоценоз — однородный участок земной поверхности с определенным составом живых организмов и косных компонентов, объединенных обменом веществ и энергии в единый природный комплекс. Биосфера — оболочка Земли, населенная живыми организмами. Буферы – вещества, которые, находясь в растворе, предотвращают резкие сдвиги рН, возникающие при добавлении кислот или оснований. Вирус – мельчайший инфекционный агент, состоящий из нуклеиновой кислоты, заключенной в белковую оболочку; может размножаться и мутировать внутри клетки-хозяина. Витамины – органические соединения, которые необходимы данному организму в малых количествах для нормального обмена веществ и должны содержаться в пище, так как организм не способен синтезировать их. Водородная связь – слабая связь между двумя молекулами (или частями одной молекулы) через атом водорода, который соединяется с двумя атомами (обычно один из них – кислород); такие связи играют важнейшую роль в структуре нуклеиновых кислот. Гамета — гаплоидная половая клетка. Гаметическая редукция хромосом — мейоз происходит при образовании гамет. Гаплоидный — содержащий одинарный хромосомный набор в клетках. Ген – биологическая единица наследственной информации, способная к само воспроизведению и рапсоложенная в определенном участке (локусе) определенной хромосомы. Гермафродит — двуполый организм, способный производить как яйцеклетки, так и сперматозоиды. Гетеротроф — питающийся готовыми органическими веществами. Гликолиз – превращение сахаров в более простые соединения в процессе обмена веществ. Гомологичные (органы) — сходные по происхождению, но не обязательно сходные по функциям. Дезоксирибоза – сахар с пятью углеродными атомами в молекуле, отличающийся от рибозы отсутствием одного из атомов кислорода; входит в состав ДНК. Денатурация – изменение физических свойств и трехмерной структуры белка, нуклеиновой кислоты или иного макромолекулярного вещества в результате какого-либо слабого воздействия, не разрушающего первичной структуры. ДНК (дезоксирибонуклеиновая кислота) – вещество, из которого состоят хромосомы; содержит генетическую информацию, закодированную в специфических последовательностях нуклеотидов, из которых построена ДНК. Диплоидия — двойной хромосомный набор в клетках. Живорождение — развитие яиц в теле самки с последующим рождением молоди. Зигота — клетка, возникающая при слиянии гамет, имеет диплоидный набор хромосом. Зиготическая редукция хромосом — мейоз происходит на фазе зиготы. Жизненный цикл — морфогенез вида между двумя одноименным фазами его циклического развития (от зиготы до зиготы и т. п.). Изогамия — образование одинаковых гамет у особей одного вида. Иммиграция — образование энтодермы путем погружения клеток и бластодермы в бластоцель. Инвагинация — образование гаструлы путем впячивания бластодермы на вегетативном полюсе. Конъюгация — половой процесс у инфузорий, сопровождающийся обменом ядерного материала между особями. Онтогенез — индивидуальное развитие организма. Орган — структурно-функциональная единица тела многоклеточного организма, образованная одной или несколькими тканями. Органелла — структурно-функциональная единица одноклеточного организма. Паразит — организм, постоянно или временно обитающий внутри другого или на нем и приносящий ему ущерб. Партеногенез — форма размножения, при которой новая особь развивается из яйцеклетки без оплодотворения. Пиноцитоз — поглощение клеткой мелких капель жидкости. Плазмодий — многоядерная амебоидная клетка. Планктон — организмы, обитающие в толще воды и не способные плыть против течения. Плейстон — полупогруженные в воду плавающие организмы. Половое размножение — форма размножения, которая сопровождается образованием гамет и последующим слиянием гамет во время оплодотворения. Почкование — бесполое размножение путем образования новых особей из выростов тела родителей. Прокариоты — организмы с клетками без ядер и органелл, окруженных мембранами. Прямое развитие — без образования стадии личинки. Радиальная симметрия — симметрия по отношению к любой плоскости, проходящей через продольную ось тела. Регенерация — восстановление утраченных частей тела организма за счет роста тканей. Реснички — двигательные органеллы клеток, по строению похожие на жгутики у жгутинокосцев. Симбиоз — взаимополезное сожительство организмов разных видов. Синцитий — многоклеточная структура с отсутствующими границами между клетками. Сперматозоид — мужская гамета, обычно способная к движению. Сперматофор — пакет сперматозоидов с защитной оболочкой. Спора — фаза в жизненном цикле некоторых паразитических простейших, выполняющая функцию расселения вида во внешней среде и содержащая молодые стадии паразита. Спорогония — бесполое размножение на фазе зиготы у споровиков с образование спорозоитов. Фагоцитоз — поглощение пищевых частиц клеткой при помощи псевдоподий. Филогенез — историческое развитие таксонов. Эволюция — происхождение и изменение живого в историческом масштабе. Эктодерма — наружный зародышевый слой, покрывающий гаструлу. Эмбрион — зародыш. Энтодерма — внутренний зародышевый листок, образующий эмбриональную кишку (гастроцель) на стадии гаструлы. Эукариоты — организмы, в клетках которых имеются окруженные мембранами ядро и органеллы. Ювенильный —неполовозрелый. Яйцеклетка — женская гамета. Яйцо — яйцеклетка или зигота, окруженная оболочками, как начальная стадия развития организма. Сложное яйцо может содержать кроме яйцеклетки желточные клетки.
РАЗДЕЛ 2. Краткий конспект лекций: Эволюция клетки Все живые существа состоят из клеток - маленьких, окруженных мембраной полостей, заполненных концентрированным водным раствором химических веществ. Простейшие формы жизни - это одиночные клетки, размножающиеся делением. Более высокоразвитые организмы, такие как мы сами, можно сравнить с клеточными городами, в которых специализированные функции осуществляют группы клеток, в свою очередь связанные между собой сложными системами коммуникаций. В известном смысле клетки находятся на полпути между молекулами и человеком. Мы изучаем клетки, чтобы понять, каково их молекулярное строение, с одной стороны, и чтобы выяснить, как они взаимодействуют для образования столь сложного организма, как человек, - с другой. Считается, что все организмы и все составляющие их клетки произошли эволюционным путем от общей предковой клетки. Два основных процесса эволюции - это 1) случайные изменения генетической информации, передаваемой от организма к его потомкам, и 2) отбор генетической информации, способствующей выживанию и размножению своих носителей. Эволюционная теория является центральным принципом биологии, позволяющим нам осмыслить ошеломляющее разнообразие живого мира. Эта глава, как и книга в целом, посвящена развитию - от молекул до многоклеточных организмов. В ней обсуждается эволюция клетки, сначала как самовоспроизводящейся единицы, состоящей из более мелких частей, а затем как строительного блока для более крупных структур. По мере изложения материала мы будем последовательно знакомиться с компонентами и функциями клетки, которые детально рассматриваются в следующих главах в основном в том же порядке. Мы узнаем, как свойства больших молекул определенного типа обеспечивают передачу потомству и выражение в фенотипе (экспрессию) наследственной информации, обусловливая эволюционный процесс. Эти молекулы, заключенные в мембрану, составляют сущность самореплицирующейся клетки. Затем мы опишем основные этапы эволюции - от небольших, похожих на бактерии клеток до значительно больших и более сложно устроенных, таких, например, как клетки современных растений и животных. Наконец, будут высказаны гипотезы о том, каким образом отдельные свободноживущие клетки породили большие многоклеточные организмы, как клетки специализировались и как, объединившись, образовали столь сложные органы, как мозг. Естественно, в эволюционном подходе есть свои опасности: большие пробелы в наших знаниях мы заполняем рассуждениями, детали которых могут быть ошибочными. Не в наших силах вернуться в прошлое и стать свидетелями уникальных молекулярных событий, происходивших миллиарды лет назад. Однако эти древние события оставили много следов, которые мы можем анализировать. Предковые растения, животные и даже бактерии сохранились как ископаемые. Но, что еще более важно, каждый современный организм содержит информацию о признаках живых организмов в прошлом. В частности, существующие ныне биологические молекулы позволяют судить об эволюционном пути, демонстрируя фундаментальное сходство между наиболее далекими живыми организмами и выявляя некоторые различия между ними. Анализируя молекулярное подобие и различие, мы пытаемся воссоздать признаки живших некогда существ. Эту задачу можно сравнить с той, которую решает ученый филолог, восстанавливая текст древнего автора, искаженный при неоднократных копированиях и редактированиях. Задача трудна и доказательства несовершенны, и все-таки этот путь дает возможность делать разумные предположения относительно основных стадий в эволюции живых клеток.
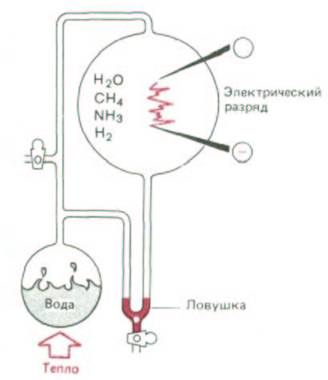
Рис. 1-1. Типичный опыт, имитирующий первозданные условия на Земле. Воду нагревают в герметичном сосуде, содержащем СН4, NH3 и Н2. Через смесь газов и водяного пара пропускают электрический разряд. Органические соединения накапливаются в U-образной ловушке.
Рис. 1-2. Некоторые соединения, которые могли образоваться в опыте, описанном на рис. 1-1. Соединения, выделенные цветом, являются важными компонентами современных живых клеток.
Задача молекулярной биологии – познание и истолкование строения, функций и взаимодействий молекул, лежащих в основе жизненных процессов. Поскольку основу жизнедеятельности определяют огромные молекулы биологических полимеров, прежде всего белков, нуклеиновых кислот и их комплексов, то их изучение и является главным направлением молекулярной биологии. Возникнув главным образом из биохимии, молекулярная биология соединяет в себе подходы и методы физики, химии, генетики, цитологии и ряда других смежных наук. В наши дни молекулярная биология заняла ключевые позиции среди биологических наук. Благодаря развитию молекулярной биологии открываются новые пути в изучении наследственных болезней человека и природы развития злокачественных новообразований. Молекулярная биология позволяет найти подходы к изысканию неизвестных эффективных лекарственных веществ, познать механизмы их действия, открывает новейшие способы борьбы со многими вирусными заболеваниями. Благодаря развитию молекулярной биологии открываются перспективы переделки наследственной природы сельскохозяйственных растений и животных. Решение задач по молекулярной биологии способствует более глубокому пониманию и прочному усвоению важнейших положений теории, наглядно иллюстрирует многообразие её практических применений, значительно повышает интерес к курсу. Задачи помогают выявить и раскрыть в теории самое главное, активно овладеть им и сознательно строить дальнейшее на этой прочной основе. Подробное изложение хода рассуждения и способов решения задач способствует выработке у студентов генетического образа мышления на основе знания основных законов и явлений наследственности.
Заключение Живые клетки скорее всего появились на Земле приблизительно 3,5 млрд, лет назад в результате спонтанной агрегации молекул. Изучение современных организмов и содержащихся в них молекул позволяет предполагать, что развитие автокаталитических механизмов, присущих живым системам, началось с эволюции группы молекул РНК, которые могли катализировать собственную репликацию. Со временем одна из этих групп согласованно катализирующих РНК приобрела способность к прямому синтезу полипептидов. Первые клетки, по-видимому, широко использовали каталитические функции и РНК, и белков, а в качестве вещества наследственности содержали только РНК. После того как накопление дополнительных каталитических белков сделало возможным развитие более эффективных и сложных клеток, двухцепочечная ДНК заменила РНК в роли хранителя генетической информации.
Лекция №2. Макромолекулы Основные клеточные макромолекулы – белки, нуклеиновые кислоты и полисахариды – это полимеры аминокислот, нуклеотидов и сахаров, синтезируемые из соответствующих малых молекул. Макромолекулы отвечают за сборку клеточных компонентов, за катализ химических реакций, за осуществление движений клетки и (самое важное) за наследственность. Выполнение всех этих жизненно важных функций обеспечивается информационным содержанием биологических макромолекул. На долю макромолекул приходится большая часть сухого вещества клетки. Их молекулярные массы составляют от 10тыс. до 1млн., т.е. по такие молекулы занимают промежуточное положение между органическими малыми молекулами и такими надмолекулярными структурами, как рибосомы или вирусы. Обычно в построении каждой макромолекулы участвуют лишь субъединицы одного семейства. Так, аминокислоты, связываясь с другими аминокислотами, образуют белки; нуклеотиды, связываясь с другими нуклеотидами, образуют нуклеиновые кислоты; а сахара, связываясь с другими сахарами, образуют полисахариды. Поскольку для нормального функционирования макромолекулы решающее значение имеет точная последовательность субъединиц (мономеров), при биосинтезе макромолекул должны действовать механизмы, точно определяющие положение каждого мономера в цепи полимера. Макромолекулярные цепи образуются с помощью ковалентных связей, которые достаточно прочны, чтобы поддерживать последовательность субъединиц макромолекулы в течение длительного времени. Но заключённая в этой последовательности информация выражается с помощью более слабых нековалентных связей. Такие слабые связи могут возникать как между разными частями одной и той же молекулы, так и между разными макромолекулами. В совокупности эти связи определяют и пространственную структуру макромолекулярных цепей, и их взаимодействие. Нековалентные связи в биологических молекулах подразделяют на три типа: ионные взаимодействия, водородные связи и вандерваальсовы взаимодействия. Ионные взаимодействия образуются между полностью заряженными группами или между частично заряженными группами. В отсутствие воды ионные силы очень велики. Они обусловливают прочность многих минералов, например мрамора и агата. В результате взаимодействия с молекулами воды заряженные группы экранируются, поэтому в водном растворе ионные взаимодействия сравнительно слабы (энергия равна примерно энергии водородной связи). Водородную связь образует атом водорода, «поделённый» между двумя электроотрицательными атомами (например, О и N). Вандерваальсовы притяжения возникают из-за флюктуаций электрических полей между любыми двумя атомами на очень близких расстояниях. Каждый атом имеет характерный вандерваальсов радиус (Н – 1,2Å; О – 2,0Å; О – 1,4Å; N – 1,5Å). Индивидуальные вандерваальсовы взаимодействия значительно слабы, но могут оказаться существенными при очень тесном сближении поверхностей двух макромолекул. Ещё одно важное слабое взаимодействие создаётся пространственной структурой воды, которая стремится вытеснить гидрофобные группы, нарушающие сеть из связанных водородными связями молекул воды, и таким образом объединяет эти гидрофобные группы и сводит к минимуму их влияние (рис.3). Такое выталкивание из водного раствора иногда считают четвёртым типом слабой нековалентной связи и называют гидрофобным взаимодействием. В водном растворе нековалентные связи примерно в 100 раз слабее ковалентных – их энергия лишь ненамного превышает среднюю энергию столкновений молекул, обусловленную тепловым движением, при 37°С. Нековалентные связи обусловливают специфичность биологического узнавания, например узнавание ферментом своего субстрата.
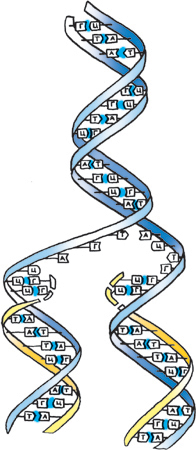
Нуклеиновые кислоты Главные функции нуклеиновых кислот состоят в хранении и передаче генетической информации. Неизвестны существа, где эти функции осуществлялись бы без их участия. Решающим фактором при отборе для этих целей в процессе эволюции именно нуклеиновых кислот явилось сочетание возможностей, во-первых, к образованию ими гигантских устойчивых линейных полимеров и, во-вторых, к сборке на матрице такого полимера его «негативной копии», которая в свою очередь может служить матрицей для синтеза точной копии исходного. Другие биополимеры не обладают указанным сочетанием свойств. Вместе с тем не стоит забывать, что непременными участниками процесса воспроизведения генетической информации являются белки. Нуклеиновые кислоты являются важнейшими регуляторами и участниками многих процессов биосинтеза. Правда, и эта их роль не уникальна – они делят её опять-таки с белками. Хранение и передача генетической информации – это одна из форм биологической памяти. У большинства живых существ хранителями наследственной информации являются наиболее консервативные по структуре и свойствам двухцепочечные дезоксирибонуклеиновые кислоты – ДНК. Лишь у части наиболее просто организованных живых существ – вирусов генетическими нуклеиновыми кислотами (ГНК) являются двухцепочечные рибонуклеиновые кислоты – РНК, а также одноцепочечные ДНК и РНК. Эволюционно РНК возникла раньше ДНК, на какой-то более поздней стадии эволюционного процесса ДНК заменила РНК в качестве вещества наследственности. ДНК представляет собой длинный неразветвлённый полимер, состоящий всего из четырёх субъединиц – дезоксирибонуклеотидов, азотистые основания которых представлены аденином (А), цитозином (Ц), гуанином (Г) и тимином (Т). Нуклеотиды связаны между собой ковалентными фосфодиэфирными связями, соединяющими 5′-атом углерода одного остатка дезоксирибозы с 3′-атомом углерода следующего остатка (рис 16). В 1953г. Джеймсом Уотсоном и Фрэнсисом Криком была построена модель структуры ДНК. Уотсон и Крик показали, что ДНК состоит из двух полинуклеотидных цепей. Каждая цепь закручена в спираль вправо, и обе они свиты вместе, т.е. закручены вправо вокруг одной и той же оси, образуя двойную спираль. Цепи антипараллельны, т.е. направлены в противоположные стороны, так что 3′-конец одной цепи располагается напротив 5′-конца другой (рис.18). Каждая цепь состоит из сахарофосфатного остова, вдоль которого перпендикулярно длинной оси двойной спирали располагаются основания. Основная особенность этой модели заключается в том, что все основания ДНК расположены внутри двойной спирали, а сахарофосфатный остов – снаружи. Отсюда следует, что основания одной цепи должны быть очень сильно сближены с основаниями другой цепи. Согласно этой модели, оптимальное соответствие цепей достигается в том случае, если при комплементарном спаривании одно большое пуриновое основание (А или Г – каждое с двойным гетероциклом) связывается с меньшим по размеру пиримидиновым основанием (Т или Ц – с одинарным гетероциклом). В противном случае условия для спаривания оснований, входящих в состав сдвоенных полинуклеотидных цепочек, были бы неблагоприятными (двум пуриновым основаниям было бы слишком «тесно», а два пиримидиновых были бы расположены в среднем слишком далеко друг от друга). Более того, построение моделей показало, что между Г и Ц или между А и Т образуется больше эффективных водородных связей, чем между любыми другими комбинациями нуклеотидов.
Репликация ДНК В комплементарности оснований ДНК заложена и возможность её удвоения, заключающаяся в разъединении комплементарных цепей и достройке к каждой из них новых комплементарных цепей (рис.23). Процесс удвоения молекулы ДНК называется репликацией (редупликацией).
Рисунок 23 Репликация двойной спирали ДНК.
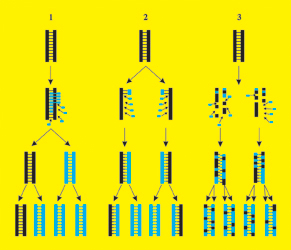
Способ воспроизведения ДНК, предложенный Уотсоном и Криком, называется полуконсервативным механизмом редупликации. Как отметил Г.Стент, можно выделить три способа редупликации: 1) консервативный – вновь синтезированная молекула не содержит материала родительской ДНК; 2) полуконсервативный – каждая вновь синтезированная молекула представлена одной родительской и одной вновь синтезированной цепями; 3) дисперсный – материал исходной молекулы случайно распределяется в обеих цепях дочерней двойной спирали (рис.24). Экспериментальные доказательства полуконсервативности редупликации ДНК были получены М.Мезельсоном и Ф.Сталем в 1957г.
Рисунок 24 Возможные пути воспроизведения ДНК. 1 – консервативный; 2 – полуконсервативный; 3 – дисперсный. Светлым показаны вновь реплицированные цепи или их участки.
Хотя принцип репликации генов прост и элегантен, реальный клеточный аппарат копирования сложен и включает в себя много различных белков. Основная реакция показана на рис. 25. Фермент ДНК-полимераза катализирует присоединение дезоксирибонуклеотида к 3′-концу цепи ДНК. Каждый нуклеотид вступает в реакцию в форме дезоксирибонуклеозидтрифосфата; высвобождение из этой активированной формы пирофосфата и его последующий гидролиз обеспечивают энергией реакцию репликации ДНК и делают её фактически необратимой. Репликация ДНК начинается с локального разделения двух комплементарных цепей. Затем каждая цепь используется в качестве матрицы для образования новой молекулы ДНК путём последовательного присоединения дезоксирибонуклеотидов. Выбор каждого следующего нуклеотида происходит на основе его способности образовывать комплементарную пару с очередным нуклеотидом родительской матричной цепи. Рост полинуклеотидной цепочки идёт только с её 3′-конца, т.е. в направлении 5′→3′. ДНК-полимераза неспособна начать матричный синтез на одноцепочечной ДНК, если нет хотя бы олигонуклеотидного биспирального участка или, иначе говоря, затравочного олигонуклеотида, комплементарного матрице. В сложных системах in vitro и in vivo всегда, по-видимому. В среде встречаются продукты распада ДНК и в их числе хотя бы незначительные количества олигонуклеотидов (три- и тетрануклеотидов, как минимум), комплементарных тому или иному участку одноцепочечной ДНК. Присоединяясь к одноцепочечной матрице, они могут служить затравкой. В последнее время установлено, что in vivo во многих случаях затравочным олигонуклеотидом является не ДНК, а РНК. Это можно объяснить тем, что в отличие от ДНК матричный синтез РНК может начинаться без затравки и, более того, РНК-полимеразы в отличие от ДНК-полимераз обладают большей специфичностью в отношении выбора начальной точки участка, подлежащего репликации. Реакция репликации ДНК требует для своего протекания присутствия двухвалентных катионов Мg2+ в концентрации 102–10-3. Она стимулируется также физиологическими значениями концентрации КСl и Н-ионов. В оптимальных условиях одна молекула энзима обеспечивает включение в строящуюся ДНК около 1000 нуклеотидов в минуту. ДНК-полимераза имеет один центр связывания нуклеозидтрифосфата, общий для всех четырёх дНТФ. Выбор из среды дНТФ, основание которого комплементарно очередному основанию матрицы, протекает без ошибок только в том случае, когда структура центра связывания субстрата не искажена. Радиоавтографические исследования, проведённые в начале 1960-х годов на целых реплицирующихся хромосомах, меченных импульсной меткой (3Н-тимидин), выявили существование ограниченной зоны репликации, перемещающейся вдоль родительской спирали ДНК. Эта активная зона из-за своей Y-образной формы была названа репликационной вилкой. В каждом таком участке синтезируется ДНК двух новых дочерних спиралей. Репликационная вилка асимметрична. Из двух синтезируемых дочерних цепей ДНК одна строится непрерывно, а другая с перерывами. Первую называют ведущей (или лидирующей), поскольку она синтезируется быстрее, а вторую – отстающей. Синтез второй цепи идёт медленнее, потому что, хотя в целом вся эта цепь строится в направлении 3′→5′, каждый из её фрагментов в отдельности наращивается в направлении 5′→3′. Благодаря тому, что ДНК на отстающей части вилки строится при помощи механизма, работающего прерывисто по типу «шитья назад иголкой», в репликационной вилке не требуется никакого другого фермента, кроме (5′→3′)-ДНК-полимеразы.
Рисунок 25 Механизм репликации ДНК (по Албертс Б. и др.,1986) Точность копирования при репликации ДНК столь велика, что в среднем на каждые 1×109 комплементарных пар, образующихся в процессе воспроизведения генома млекопитающих, насчитывающего 3×109 пар оснований, приходится лишь около одной ошибки. Процесс репликации ДНК требует совместного действия многих белков. В нём участвуют: 1) ДНК-полимераза и РНК-праймаза, катализирующие полимеризацию нуклеозидтрифосфатов; 2) ДНК-геликазы и дестабилизирующие белки, помогающие расплести спираль ДНК, которую предстоит копировать; 3) ДНК-лигазы и фермент, разрушающий молекулы РНК-затравки; они нужны для сшивания прерывисто синтезируемых фрагментов отстающей цепи ДНК; 4) ДНК-топоизомеразы, помогающие решить «проблему кручения» спирали ДНК; 5) инициаторные белки, способствующие образованию новой репликационной вилки в точке начала репликации.
Лекция №3. Молекулярные механизмы транскрипции. Матричный синтез РНК.
Цель лекции: Ознакомить с основными механизмами траскрипции. Лекция №4. Биосинтез белка. Трансляция генетической информации. Синтез белка. Трансляция Транспортные РНК
Роль главных агентов в процессе синтеза белка играют молекулы тРНК: к ним присоединяются аминокислоты перед полимеризацией, т.е. перед объединением в полипептиды. Присоединяясь к молекуле тРНК своим карбоксильным концом, аминокислоты активируются – переходят в богатую энергией форму, способную спонтанно образовать пептидную связь, что и приводит к синтезу полипептидов. Этот процесс активации – необходимый этап белкового синтеза, поскольку свободные аминокислоты не могут прямо присоединяться к полипептидной цепи. (Спонтанно способен идти лишь обратный процесс – гидролитический разрыв пептидных связей.) В каком именно месте будет присоединена к растущей полипептидной цепи данная аминокислота, зависит не от самой аминокислоты, а от присоединившей её молекулы тРНК. Существует специальный набор ферментов, так называемых аминоацил-тРНК-синтетаз, которые присоединяют аминокислоты к соответствующим молекулам тРНК. Для каждой из аминокислот имеется своя особая синтетаза. Реакция протекает в два этапа и приводит к образованию молекулы аминоацил-тРНК. Реакция образования аминоацил-тРНК обратима, так как энергия, освобождающаяся при отщеплении пирофосфата, практически не растрачивается, а резервируется при формировании эфирной связи между карбонилом аминокислоты и остатком ά-ортофосфата. Молекулы тРНК играют роль конечных «адапторов», переводящих информацию, заключённую в нуклеотидной последовательности нуклеиновой кислоты, на язык белка. Транспортные РНК относятся к категории относительно низкополимерных РНК клетки. Их молекулярный вес составляет всего лишь 25 – 28 тыс. дальтон. Единственная полинуклеотидная цепочка состоит из 75 – 85 нуклеотидов. Для нуклеотидного состава тРНК характерно преобладание гуанина и цитозина (суммарная молярная доля – около 60%). Структура и свойства тРНК, во-первых, обеспечивают связывание остатка аминокислоты в форме, обладающей достаточным запасом энергии для последующего образования пептидной связи; во-вторых, определяют точную ориентацию аминокислотного остатка на рибосоме и, в-третьих, обеспечивают строгую специфичность и выбора, и включения в строящийся пептид транспортируемой аминокислоты. Первая функция сама по себе не требует специфичности взаимодействия с активированными аминокислотами, и ей соответствует универсальный для всех тРНК 3′-концевой участок молекулы, так называемый акцепторный, который включает тринуклеотид ЦЦА. Именно 3′-ОН-группа концевого аденина замещается остатком аминокислоты. Вторая функция требует жёсткости и ряда общих особенностей структуры всех тРНК. Высокая доля спирализованных участков, в которых преобладают наиболее прочные пары – ГЦ, обеспечивает необходимую жёсткость вторичной структуры молекулы (рис.27). В молекулах тРНК имеются четыре спирализованных участка (рис.27, А, Б, В, Г), три из которых (рис.27, Б, В, Г) увенчаны петлями из неспаренных нуклеотидов, и, кроме того, имеется центральная неспирализованная область. 3′- и 5′-концы полинуклеотидной цепочки объединены в наиболее значительный спирализованный участок (7 пар), завершающийся акцепторным олигонуклеотидом ЦЦА (рис.27, Д).
Рисунок 27 Схема строения тРНК
Противостоящая акцепторному концу петля содержит тринуклеотид – антикодон, который обеспечивает специфичность взаимодействия с мРНК (рис. 27, Е). Нуклеотиды, образующие антикодон, всегда расположены в середине петли. Выполнение третьей функции возможно лишь при условии существования большого числа разновидностей тРНК. Основная реакция в синтезе белка – это реакция, приводящая к образованию пептидной связи между карбоксильной группой на конце растущей полипептидной цепи и свободной аминогруппы аминокислоты. Белковая цепь синтезируется, следовательно, путём её постепенного наращивания от аминного конца к карбоксильному. На протяжении всего процесса растущий карбоксильный конец полипептидной цепи остаётся в активированном состоянии, будучи связан ковалентной связью с тРНК (в молекуле пептидил-тРНК). В процессе синтеза белка каждая добавляемая аминокислота несёт с собой энергию активации, необходимую не для её собственного присоединения, а для присоединения следующей аминокислоты. В процессе трансляции нуклеотидная последовательность мРНК считывается группами по три нуклеотида по мере того, как считывающий «аппарат» перемещается вдоль молекулы мРНК в направлении 5′→3′. Каждая аминокислота соответствует определённому триплету нуклеотидов (кодону) в молекуле мРНК, который спаривается с последовательностью из трёх комплементарных нуклеотидов в антикодоновой петле молекулы тРНК. Выбор аминокислоты, присоединяемой в каждый данный момент к растущему концу полипептидной цепи, определяется кодоном.
Лекция №5. Структура рибосом и трансляция.
Цель лекции: Ознакомить с ультраструктурой рибосом. Изучить основные этапы трансляции.
Структура рибосом Рибосомы найдены во всех известных живых существах, кроме истинных вирусов и некоторых других особенно просто устроенных клеточных паразитов, которые в своём развитии используют рибосомы клетки-хозяина. Рибосомы эукариот и прокариот сходны по своей структуре и функции. Каждая из них состоит из двух субчастиц – большой и малой, обратимо диссоциирующих после завершения синтеза одного полипептида (рис.28). В эукариотических рибосомах приблизительно половину их массы составляет РНК; малая субчастица состоит из одной молекулы рибосомной РНК (рРНК), связанной приблизитель
|
||||
|
|
Последнее изменение этой страницы: 2016-12-28; просмотров: 223; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.191.67.90 (0.014 с.) |