
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Общие принципы регуляции двигательной деятельностиСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте Теория функциональной системы. Современные представления об организации целостных актов поведения живых организмов отражены в теории функциональной системы (П. К. Анохин). Для удовлетворения своих потребностей в условиях постоянного изменения внешней среды организму необходимо ставить перед собой определенные задачи и в своей поведенческой деятельности добиваться достижения намеченного результата. Согласно теории П. К. Анохина, именно полезный результат является решающим фактором поведения и для его достижения в нервной системе формируется группа взаимосвязанных нейронов — так называемая функциональная система. Сколько нервных клеток из огромного их числа будет включено в эту систему, какой уровень активности этих клеток необходим в данный момент, какие взаимосвязи должны возникнуть между ними и какие возможные взаимосвязи должны быть исключены — все это определяется намечаемым результатом. Следовательно, важная задача деятельности системы — выявление и оценка результата действия. Организм в своем поведении использует далеко не все структурные и функциональные свои возможности (степени свободы), а оставляет лишь необходимые. Без этого деятельность его была бы хаотичной, нецелесообразной. Создание функциональной системы необходимо для того, чтобы отдельные ее элементы действовали не самостоятельно и независимо друг от друга, а подчинялись общей задаче — получить искомый результат. Деятельность функциональной системы можно разбить на отдельные последовательные этапы: 1) обработка всех сигналов поступающих из внешней и внутренней среды организма — так называемый афферентный синтез; 2) принятие решения; 3) создание (на основе принятого решения) представления об ожидаемом результате действия и формирование конкретной программы действий для достижения этого результата; 4) анализ полученного результата и уточнение программы действий. На первом этапе (афферентный синтез) в нервных центрах одновременно взаимодействуют 4 типа поступающих к ним раздражении: 1) пусковая афферентация—сигналы, вызывающие действие; 2) обстановочная афферентация — все остальные внешние раздражения, создающие афферентный фон; 3) мотивация — собственные потребности организма, доминирующие в данный момент: биологические мотивации — жажда, голод, страх и другие, а также высшие мотивации, формирующиеся у человека под влиянием социальных факторов (желание добиться какой-либо цели в жизни, достичь определенного результата в спорте и др.); 4) память—имеющиеся в нервной системе следы прежних раздражении, накопленный опыт. Все эти виды сигналов должны одновременно обрабатываться в нервных клетках и сопоставляться друг с другом. При оценке текущих афферентных раздражении производится их сопоставление со следами прежних раздражении, хранящихся в памяти. Большую роль в отборе сигналов играют доминирующие мотивации, так как они повышают восприимчивость организма к наиболее важным в данный момент сигналам. На основании афферентного синтеза принимается решение («что надо делать») и вырабатывается программа действий («как надо делать»). При этом производится оценка всех возможных путей решения возникших перед организмом задач и ответных реакций организма и выбирается лишь одно необходимое решение. Одновременно в мозгу создается представление о том, какой результат должен быть получен. Динамическая функциональная система — это организованная система, результат действия которой сообщается в центральную нервную систему. Без постоянной оценки результата действия невозможно выполнить задуманную программу, вносить в нее поправки для улучшения результата. Например, совершенствование техники выполнения упражнения невозможно без постоянной информации о его результатах. Представление о функциональной системе П. К. Анохина имеет универсальное значение, так как помогает понять различные стороны деятельности организма (механизмы его поведения, закономерности формирования различных функций в процессе индивидуального развития, особенности их нарушения при патологии и возможные пути восстановления). Циклическая многоуровневая система регуляции движений. Выполнение двигательных актов осуществляется обширным комплексом нейронов, расположенных в различных отделах центральной нервной системы. Каждый из нервных центров, входящих в данный комплекс, может воздействовать как на нижележащие, так и на вышележащие центры. При этом между ними могут возникать циклы взаимных влияний от высших центров к низшим и обратно. Например, очень часто наблюдается циркуляция импульсов между корой и промежуточным мозгом: импульсы из коры больших полушарий могут влиять на активность нейронов промежуточного мозга, а те, в свою очередь, могут воздействовать на корковые нейроны. В каждом подобном цикле роль задающих звеньев принадлежит преимущественно вышележащим центрам, а активность нижележащих центров является регулируемым звеном. Эта активность поддерживается на заданном уровне в зависимости от потребностей организма. Наличие циклических взаимосвязей между различными отделами нервной системы позволяет говорить о замкнутой системе регулирования, или замкнутом цикле регулирования. В процессе двигательной деятельности одновременно возникает множество замкнутых циклов регулирования (например, между корой и ретикулярной формацией, между корой и мозжечком, между мотонейронами спинного мозга и мышцей и др.). Таким образом, система регуляции движения является многоэтажной (многоуровневой) системой. Важную роль в этой сложной функциональной системе играет кора больших полушарий, которая управляет процессами, протекающими в этих циклах регулирования, и направляет их на решение общей задачи—достижение рабочего эффекта, или цели движения. Программирование. Для решения поставленных задач в центральной нервной системе вырабатываются программы двигательных действий. Они определяют набор и последовательность включения двигательных актов. В соответствии с этой программой включаются команды к эфферентным нейронам и далее—команды к мышцам. Программы двигательных действий у человека разнообразны. Они обеспечивают бытовую, производственную и спортивную его деятельность. При этом команды к действию (например, выстрел стартового пистолета, команда «марш!») являются лишь сигналами о начале работы, сама же мышечная работа развертывается по имеющейся программе. В ряде случаев программирование протекает в условиях крайнего дефицита времени, (например при спортивных играх, фехтовании, борьбе и др.). В этих случаях требуется высокая скорость переработки поступающей информации и выполнения ответных действий.
Обратные связи. Для функциональной системы регуляции движений недостаточно лишь наличия программирующих и управляющих нервных центров. Характерной особенностью подобных систем регуляции является наличие обратной афферентации, или так называемой обратной связи, от рабочего органа к регулирующим нервным центрам (рис. 57). Осведомление центральной нервной системы о деятельности, вызванной на периферии ее управляющими импульсами, происходит при помощи афферентных сигналов, отправляемых органами зрения проприорецепторами мышечной суставного аппарата и другими рецепторами. Таким образом, с помощью обратных связей производится оценка выполненных движений, определяется их эффективность. Аппараты сравнения. Эти нервные аппараты, получая обратную сигнализацию, сличают заданную программу с реальными результатами ее выполнения (П. К. Анохин, Н. А. Бернштейн и др.). Имеются данные, что такие аппараты находятся в лобных долях, подкорковых ядрах (хвостатые тела) и других образованиях. Если есть расхождения фактического и требуемого действия, то в аппаратах сравнения возникают сигналы об ошибках—так называемые импульсы рассогласования. Соответственно этим импульсам в систему, управляющую движением, вносятся необходимые поправки — коррекции. В результате этого в процессе выполнения движения можно сохранить его основные черты, целесообразность, несмотря на изменение условий его осуществления. Так, сохраняется, например, индивидуальный характер походки человека при различиях в обуви, грунте, скорости движения, положении центра тяжести и т. д. Позно-тонические реакции Общее представление о двигательной деятельности человека. Двигательная деятельность включает в себя процессы осуществления двигательных актов и процессы поддержания позы. В основе двигательной деятельности человека лежат 2 формы механической реакции мышечных волокон: 1) их фазное (тетаническое) сокращение и 2) длительное тоническое напряжение. Фазная деятельность обеспечивает быстрые и сильные сокращения мышц при выполнении движений, а также коррекции позы. Эти сокращения осуществляются быстрыми двигательными единицами. Тоническое напряжение отличается от фазного медленным развитием, большей экономичностью, преобладанием изометрического режима и преимущественным участием медленных двигательных единиц. Оно лежит в основе поддержания определенной позы тела. Если при поддержании позы требуется значительное усилие мышц (например для выполнении угла в упоре), то включаются фазные сокращения мышц. Процессы поддержания позы и двигательные акты скелетных мышц тесно связаны между собой. С одной стороны, движения всегда происходят на фоне определенной позы тела; с другой стороны, процессы поддержания позы обеспечиваются не только тоническим напряжением мышц, но и их фазной деятельностью. В двигательной деятельности человека различают произвольные и непроизвольные движения и позы. Произвольные движения вырабатываются с участием сознания, а по мере их освоения могут происходить и бессознательно. Ведущую роль в осуществлении произвольных движений играют высшие интегративные области коры больших полушарий (лобные доли и др.). Еще до их выполнения в мозгу создается модель предстоящего движения, его программа. Человек сознает цель этого движения или необходимость принятия определенной позы. В процессе выполнения двигательного акта он, внося коррекции в его выполнение, добивается запрограммированного результата действия. Непроизвольные движения и позы, выполняемые бессознательно, являются преимущественно безусловными рефлексами (ориентировочными, защитными и др.). Непроизвольными также становятся те произвольные движения и позы, которые автоматизируются при многократном их повторении. Непроизвольная двигательная деятельность может осуществляться различными отделами нервной системы. Рефлекторная природа мышечного тонуса. В поддержании тонического напряжения мышц участвует сложная многоуровневая система нервных центров, находящаяся под контролем коры. больших полушарий. Многочисленные опыты на животных, а также наблюдения за нарушениями мышечной деятельности у человека в клинике позволили выявить значение отдельных участков мозга в осуществлении и регуляции мышечного тонуса.
Для возникновения тонуса в мышцах достаточна уже рефлекторная деятельность спинного мозга. Однако тонкое перераспределение тонуса между различными мышцами, необходимое для протекания реакций целостного организма, осуществляется более высокими Этажами центральной нервной системы, а его произвольная регуляция — корой больших полушарий. В наиболее простом случае появление тонуса обусловлено сокращением мышцы в ответ на раздражение проприорецепторов при ее растяжении. Такой рефлекс, осуществляемый спинным мозгом называется миотатическим рефлексом на растяжение. Если это растяжение длительно, то рефлекторное сокращение носит тоже длительный тонический характер. При растяжении мышцы возбуждаются чувствительные нервные окончания в мышечных веретенах. Импульсы от них направляются по толстым афферентным волокнам в передние рога спинного мозга, где непосредственно (без участия вставочных нейронов) передаются на альфа-мотонейроны и вызывают сокращение мышц. Такая же двухнейронная рефлекторная дуга лежит в основе аналогичного сухожильного рефлекса — сокращения мышцы при ударе по ее сухожилию. К сухожильным рефлексам относят рефлекс ахиллова сухожилия, коленный рефлекс и др. В естественных условиях основным раздражителем рецепторов мышечных и сухожильных веретен является сила тяжести, растягивающая скелетные мышцы, особенно мышцы-разгибатели. Поражение спинного мозга приводит к исчезновению тонуса мышц. Гамма-регуляция мышечного тонуса. Степень тонического напряжения мышцы зависит от частоты импульсов, посылаемых к ней тоническими альфа-мотонейронами. Частота разрядов альфа-мотонейронов, в свою очередь, регулируется импульсами от проприорецепторов той же самой мышцы. Получается замкнутое кольцо между мышцей и иннервирующими ее мотонейронами. Однако потоки импульсов в этом кольце могут регулироваться вышележащими этажами нервной системы с помощью гамма-мотонейронов спинного мозга (рис. 58). Разряд гамма-мотонейронов повышает чувствительность мышечных веретен. В результате увеличивается поток импульсов от рецепторов к альфа-мотонейронам и от альфа-мотонейронов к мышце. Тем самым повышается мышечный тонус. Изменение напряжения мышц под влиянием деятельности гамма-мотонейронов называется гамма-регуляцией. Активность гамма-мотонейронов находится под контролем ретикулярной формации ствола мозга. В регуляции их деятельности, а следовательно, и в регуляции тонуса скелетных мышц участвуют мозжечок, подкорковые и другие структуры экстрапирамидной системы. Высший контроль осуществляется корой больших полушарий. Значение различных отделов головного мозга в регуляции тонуса скелетных мышц. Различные отделы головного мозга, посылая импульсы к вставочным нейронам и мотонейронам спинного мозга, могут регулировать тонус скелетных мышц, изменяя тем самым позные и двигательные реакции организма. Влияния на мышечный тонус оказывают обе системы головного мозга — как специфическая, так и неспецифическая. Неспецифическая система вызывает общее изменение тонуса различных мышц: усиление тонуса осуществляет активирующий отдел среднего мозга, а угнетение — тормозящий отдел продолговатого мозга. Специфические системы действуют избирательно — на отдельные группы мышц. Усиление тонуса мышц-сгибателей вызывают кортико-спинальная, кортико-рубро-спинальная и частично кортико-ретикуло-спинальная системы (последняя оказывает также неспецифическое диффузное влияние). Одновременно эти системы снижают тонус мышц-разгибателей. В противоположность им вестибуло-спинальная система повышает тонус мышц-разгибателей и тормозит тоническое напряжение мышц-сгибателей. Изменения тонуса могут происходить очень быстро, нередко опережая двигательные акты. Это позволяет организму подготовиться к необходимому движению, заранее изменив позу тела. В опытах на животных было показано, что при перерезке головного мозга между продолговатым и средним мозгом резко повышается тонус мышц-разгибателей. Это явление было названо децеребрационной ригидностью (децеребрация — лишение животного всех отделов головного мозга выше продолговатого, а ригидность — это твердость). После децеребрации у кошки напряжены мышцы спины, голова запрокинута назад, а хвост и конечности вытянуты. Этого не происходит в целостном организме, так как влияния из вестибулярных ядер продолговатого мозга, усиливающие тонус мышц-разгибателей, уравновешиваются в необходимой степени воздействиями других нисходящих систем, усиливающих тонус мышц-сгибателей. Правильное распределение тонуса различных мышц, которое лежит в основе организации определенной позы, обеспечивается деятельностью мозжечка, подкорковых ядер (полосатого тела и бледного ядра) и коры больших полушарий. Мозжечок не имеет прямых связей со спинным мозгом, но через красные ядра среднего мозга повышает тонус мышц-сгибателей, а через вестибулярные ядра продолговатого мозга усиливает тонус Мышц-разгибателей. Поражения мозжечка нарушают правильное перераспределение тонуса скелетных мышц, что изменяет не только позные, но и двигательные реакции организма. При мозжечковых расстройствах вследствие снижения и ненормального распределения тонуса мышц конечностей нарушается походка (атаксия). Бледное ядро угнетает тонус мышц, а при его поражениях тонус усиливается. Полосатое тело, регулируя деятельность бледного ядра, снижает его угнетающее действие. Высший контроль тонической активности мышц осуществляет кора больших полушарий, в частности ее моторные, премоторные и лобные области. С ее участием происходит выбор наиболее целесообразной для данного момента позы организма, обеспечивается ее соответствие двигательным задачам. Непосредственное отношение к регуляции тонуса имеют медленные пирамидные нейроны коры — так называемые нейроны положения, образующие медленную часть кортико-спинального пути (пирамидного тракта). По этому пути передаются прямые корковые воздействия на спинной мозг. Кроме того, изменяя активность ретикулярной формации ствола мозга, подкорковых ядер, мозжечка, ядер среднего и продолговатого мозга, кора больших полушарий оказывает через них параллельное влияние на тонус мышц. Позы. Позой называется закрепление частей скелета в определенном положении. Значение поз в двигательной деятельности человека. Организация позы необходима для преодоления силы земного притяжения. Поза человека служит для сохранения равновесия тела в состоянии двигательного покоя и при выполнении статической или динамической работы, а также для фиксации ряда суставов, необходимой для осуществления движения в других суставах. При многих видах трудовой и спортивной деятельности человеку приходится принимать новые необычные позы, без которых невозможно движение. Поза, принимаемая при работе, называется рабочей позой. Правильная ее организация способствует повышению работоспособности человека. Наконец, большое значение имеют предрабочие изменения позы, которые заранее компенсируют возможные изменения центра тяжести тела и предотвращают его падение, а также препятствуют действию реактивных сил. Реакции поддержания позы обеспечиваются чаще всего тоническим напряжением мышц, а при больших нагрузках и при коррекциях, исправляющих позу,— фазными сокращениями мышц.
Частным случаем позных реакций является поддержание заданного усилия мышцы и заданного суставного угла. Усилия мышц не сохраняются постоянными. Каждый раз, когда мышца расслабляется и возникают отклонения от заданного усилия или от определенного положения костей, появляются небольшие фазные сокращения работающих мышц, исправляющие эти отклонения. Нарастание усилия мышц достигается благодаря увеличению числа активных альфа-мотонейронов спинного мозга. Таким образом, сохранение достоянного усилия и суставного угла поддерживается как тоническими, так и фазными сокращениями мышц. Известно, что человек может (особенно в результате мышечной тренировки) очень точно дозировать усилия своих мышц и определять положение суставов. Информацию о положении конечностей сообщают в кору больших полушарий многие рецепторы — мышечные и сухожильные, зрительные, кожные, но особенно большое значение имеют рецепторы суставных, сумок. Позно-тонические реакции мышц часто бывают кратковременными, так как возникают только в определенные фазы движения. Они способствуют, в частности, фиксации суставов при отталкивании от опоры в процессе ходьбы. Электромиографические исследования ходьбы показали, что наряду с работой мышц-антагонистов, вызывающих сгибания и разгибания нижних конечностей (например, передней большеберцовой мышцы и икроножной мышцы ноги), в фазу отталкивания носком от опоры происходит тоническое напряжение мышц стопы, фиксирующих ее свод. Установочные рефлексы. Эти рефлексы способствуют сохранению позы. К ним относятся статические и статокинетические рефлексы, в осуществлении которых большое значение имеет продолговатый и средний мозг. Статические рефлексы возникают при изменении положения тела или его частей в пространстве: 1) при изменении положения головы в пространстве — это так называемые лабиринтные рефлексы. возникающие в результате раздражения рецепторов вестибулярного аппарата; 2) при изменении положения головы по отношению к туловищу — шейные рефлексы, с проприорецепторов мышц шеи и 3) при нарушении нормальной позы тела — выпрямительные рефлексы с рецепторов кожи, вестибулярного аппарата и сетчатки глаз. Например, при отклонении головы назад повышается тонус мышц-разгибателей спины, а при наклоне вперед — тонус сгибателей (лабиринтный рефлекс). Подобные реакции имеют место при выполнении многих физических упражнений. Так, выполнение гимнастом стойки на кистях облегчается при отклонении головы назад выполнение группировки во время прыжков в воду или акробатических прыжков — при наклоне головы вперед. Выпрямительные рефлексы — это последовательные сокращения мышц шеи и туловища которые обеспечивают возвращение тела в вертикальное положение теменем кверху. У человека они проявляются, например, во время ныряния. Статокинетические рефлексы компенсируют отклонения тела при ускорении или замедлении прямолинейного движения, а также при вращениях. Например, при быстром подъеме усиливается тонус сгибателей, и человек приседает, а при быстром спуске усиливается тонус разгибателей, и человек выпрямляется — это так называемый лифтный рефлекс. При вращении тела реакции противовращения проявляются в отклонении головы, тела и глаз в сторону, противоположную движению. Движение глаз со скоростью вращения тела, но в противоположную сторону и быстрое возвращение в исходное положение — нистагм глаз — обеспечивают сохранение изображения внешнего мира на сетчатке глаз и тем самым зрительную ориентацию. В двигательной деятельности человека часто возникают ситуации, когда требуется подавить эти установочные рефлексы. Произвольное подавление врожденных установочных рефлексов продолговатого и среднего мозга обеспечивается тормозящими влияниями из коры полушарий. Например, для спринтера невыгодно раннее выпрямление туловища при стартовом разбеге, поэтому корой больших полушарий тормозится выпрямительный рефлекс. Двигательная деятельность Движения человека являются комплексом различных двигательных актов, которые обеспечиваются как сравнительно простыми двигательными рефлексами, так и сложнейшими их комбинациями. Двигательные акты различной сложности. Рассматривая различные двигательные акты человека, можно выделить элементарные двигательные рефлексы, более сложные—ритмические рефлексы и, наконец, особенно сложные формы двигательной деятельности, обеспечивающие поведение человека. Элементарные двигательные рефлексы. При сложных движениях человека используются элементарные двигательные рефлексы, осуществляемые спинным мозгом. К простым безусловным двигательным рефлексам спинного мозга относятся: 1) рефлексы, на растяжение (миотатические и сухожильные рефлексы—рис. 60, А), 2) сгибательные рефлексы на раздражение кожных рецепторов (рефлексы удаления от раздражителя — см. рис. 60, Б) и 3) рефлексы отталкивания (рефлексы сближения с опорой—см. рис. 60, В).
При раздражении рецепторов кожи (болевых, температурных и др.) возникает сокращение мышц-сгибателей, которое позволяет отдернуть конечность от повреждающего раздражителя. Этот сгибательный кожный рефлекс носит защитный характер. В его основе лежит полисинаптическая рефлекторная дуга. В отличие от моносинаптического рефлекса на растяжение при сгибательном рефлексе быстрее наступает утомление; время этого рефлекса в связи с наличием вставочных нейронов значительно больше. Одна из разновидностей элементарных рефлексов спинного мозга—рефлекс отталкивания, описанный Ч. Шеррингтоном. Этот Рефлекс возникает при раздражении кожи стопы опорой. Однако, в Отличие от сгибательного рефлекса, он приводит не к отдергиванию конечности от раздражителя, а к сближению с раздражителем. В первом случае мы имеем защитный рефлекс, предохраняющий организм от разрушения. Во втором случае—биологически целесообразный акт, обеспечивающий контакт с опорой при стоянии и отталкивание от нее при передвижениях. Этот рефлекс лежит в основе сложных локомоций: ходьбы, бега, прыжков в длину ил' высоту и др. Ритмические рефлексы. Составной частью различны» сложных двигательных действий, как произвольных, так и непроизвольных, часто являются ритмические рефлексы. Они особенно выражены при выполнении циклических движений. Их возникновение и протекание связаны с проявлением механизмов взаимосочетанной (реципрокной) иннервации мышц-антагонистов. Эта форма рефлексов — одна из древних и относительно простых форм — имеет большое значение в организации многих сложных движений. Например, включение ритмических циклоидных движений в акт письма позволяет человеку перейти от отдельного начертания букв к обычной письменной скорописи; то же самое происходит при освоении акта ходьбы — переход от отдельных шагов к ритмической походке (Н. А. Бернштейн). Наиболее простая форма ритмического рефлекса — чесательный рефлекс животных, который обеспечивается попеременным сокращением и расслаблением одних и тех же мышц одной конечности. Возбуждение центров-сгибателей сопровождается одновременным торможением центров мышц-разгибателей, в чем проявляется реципрокная их иннервация. В последующей фазе движения в этих центрах возникают противоположные процессы—возбуждение вслед за торможением и торможение вслед за возбуждением. При этом проявляется так называемый рефлекс отдачи: после акта сокращения наступает расслабление, а затем снова акт сокращения. Рефлекс отдачи — это рефлекс на растяжение с проприорецепторов мышц и сухожилий, в результате которого возникает сокращение растянутого разгибателя. Такая простая форма ритмического рефлекса часто встречается в двигательной деятельности человека (например, при забивании молотком гвоздей). Более сложная форма ритмического рефлекса — шагательный рефлекс, лежащий в основе ходьбы, бега и других локомоций. Для осуществления шагательных движений требуется участие уже не одной, а двух симметричных конечностей. Подключение второй конечности к координированному двигательному акту происходит в результате осуществления перекрестного рефлекса. Он хорошо виден на спинальном животном. Раздражение рецепторов кожи вызывает сгибание раздражаемой конечности и одновременное разгибание одноименной конечности на противоположной стороне тела. Этот рефлекс осуществляется с участием вставочных тормозных нейронов, вызывающих торможение мотонейронов на противоположной стороне спинного мозга. В целостном организме человека сгибание одной ноги вызывает перекрестный разгибательный рефлекс другой ноги, принимающей на себя тяжесть тела при стоянии, ходьбе и пр. Следовательно, механизмы шагательных движений заложены уже на уровне спинного мозга. У спиналыюго животного в первой фазе движения болевое раздражение рецепторов кожи вызывает сгибательный рефлекс раздражаемой конечности (табл. 4). Одновременно при участии тормозных клеток Рэншоу происходит торможение центров мышц-разгибателей этой же конечности и торможение центров мышц-сгибателей симметричной конечности. Во вторую фазу движения возникает рефлекс отдачи с проприорецепторов растянутых мышц-разгибателей раздражаемой конечности и мышц-разгибателей другой стороны тела. Импульсы от этих рецепторов вызывают рефлекторное сокращение мышц-разгибателей и одновременное торможение центров мышц-сгибателей той же конечности и мышц разгибателей симметричной конечности. Последовательная смена этих двух фаз движения и составляет ритмический акт шагания. Включение этого спинального механизма производится с более высоких этажей нервной системы. В так называемой «локомоторной области» среднего мозга имеются центры, электрическое раздражение которых вызывает шагательные движения конечностей. В осуществлении шагательного рефлекса принимает участие мозжечок. Удаление одного его полушария у животных приводит к искажению движений (особенно передней конечности). Движения сильно варьируют по амплитуде, заметно искажаются при малейших посторонних воздействиях, происходит чрезмерное сгибание локтевого сустава в фазе переноса конечности. Таблица 4 Процессы возбуждения (+) и торможения (—) в нервных центрах спинного мозга при осуществлении шагательного рефлекса
Протекание ритмических рефлексов, в том числе шагательного, связано с деятельностью различных отделов неспецифической системы. При активирующих влияниях происходит нарастание амплитуды движений, а при угнетающих влияниях — ее снижение. Важное значение в регуляции ритмических рефлексов имеют подкорковые ядра — бледное ядро и полосатое тело, обеспечивающие их автоматическое протекание, содружественные движения конечностей, вспомогательные реакции (фиксацию суставов и др.). Высшим регулятором рефлексов является кора больших полушарий, особенно ее премоторная область. Благодаря коре ритмические движения (например, простой акт ходьбы) приобретают определенное смысловое значение, включаются как составной элемент сложные акты поведения. Об участии различных корковых областей в регуляции ритмических рефлексов свидетельствует появление на электроэнцефалограмме медленных потенциалов, синхронных по частоте с темпом движений. Протекание ритмических рефлексов облегчается тем, что между нервными центрами и работающими мышцами возникают согласованные во времени потоки нервных импульсов. Прохождение импульсов от нервных центров на периферию к рабочим органам и от мышц обратно к нервным центрам осуществляется в определенном темпе — в темпе рабочих движений. Налаживание этого темпа и стабильное его поддержание, включение древних ритмических автоматизмов в систему регуляции движений значительно упрощают управление ритмическими рефлексами, делают эту двигательную деятельность более экономичной. Сложные формы двигательной деятельности. В целостном поведении простые рефлексы, сочетаясь, обусловливают сложные двигательные действия. Социальные условия жизни человека намного усложняют его двигательную деятельность, приводя к появлению специально человеческих форм движений: бытовых, производственных, спортивных. Простые и сложные ритмические рефлексы лежат в основе циклической двигательной деятельности человека: ходьбы, бега, плавания, гребли, ходьбы на лыжах, езды на велосипеде и пр. Большое значение в их осуществлении имеют механизмы реципрокной иннервации мышц-антагонистов (рис. 61). Однако в целом ряде движений требуется одноименная работа мышц-антагонистов — в одной конечности или даже в обеих. В этих случаях реципрокные отношения, характерные для спинного мозга, подавляются центрами головного мозга. Особенно часто встречаются симметричные (а не реципрокно-перекрестные) движения в деятельности верхних конечностей человека, т. е. происходит одновременное сокращение одноименных мышц левой и правой рук. Наиболее сложные формы движений представляют собой многофазную цепь рефлексов, которая основана уже не на элементарных реципрокных отношениях, а является целостным двигательным навыком, образующимся в процессе обучения по механизму условных рефлексов.
В организации двигательного акта участвуют самые различные отделы коры больших полушарий. Моторная зона коры (поле 4) посылает импульсы к отдельным мышцам, преимущественно к дистальным мышцам конечностей. Объединение отдельных элементов движения в целостный акт осуществляют вторичные поля (6-е и 8-е) премоторной области. Они определяют последовательность двигательных актов, формируют ритмические серии движении, регулируют тонус мышц. Задняя центральная извилина коры — общечувствительная область—обеспечивает
|
||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-09-20; просмотров: 1126; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.62 (0.02 с.) |

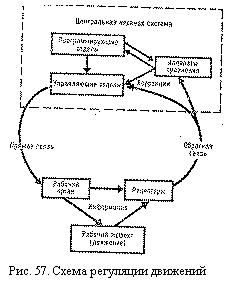 Программы двигательных действий могут быть очень сложными, особенно при нестандартной работе, так как для достижения одного и того же результата в изменяющихся условиях (например, для успешного выполнения броска в баскетбольную корзину при разных игровых ситуациях) могут потребоваться включение различных мышечных групп, различная тактика поведения.
Программы двигательных действий могут быть очень сложными, особенно при нестандартной работе, так как для достижения одного и того же результата в изменяющихся условиях (например, для успешного выполнения броска в баскетбольную корзину при разных игровых ситуациях) могут потребоваться включение различных мышечных групп, различная тактика поведения.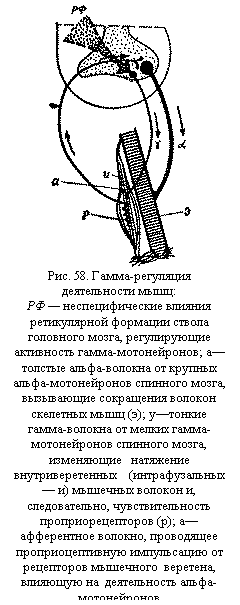 рецепторов мышечного веретена, влияющую на деятельность альфа-мотонейронов ">По своей природе мышечный тонус является рефлекторным актом.
рецепторов мышечного веретена, влияющую на деятельность альфа-мотонейронов ">По своей природе мышечный тонус является рефлекторным актом.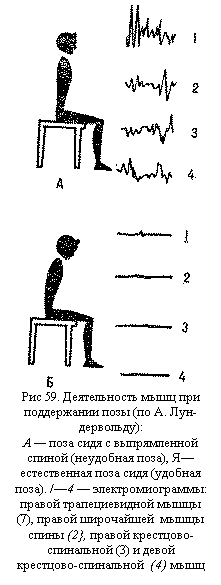 Во время отдыха или выполнения работы человек выбирает наиболее удобную позу, при которой возможно минимальное напряжение скелетных мышц (рис. 59). Организация такой оптимальной позы часто требует длительной тренировки. Например, при стрельбе из положения стоя только у высококвалифицированных стрелков тоническая активность мышц, обеспечивающих стояние, минимальна (на электромиограмме регистрируется наименьшая электрическая активность мышц). У спортсменов низкой квалификации и новичков наблюдается значительное напряжение этих мышц.
Во время отдыха или выполнения работы человек выбирает наиболее удобную позу, при которой возможно минимальное напряжение скелетных мышц (рис. 59). Организация такой оптимальной позы часто требует длительной тренировки. Например, при стрельбе из положения стоя только у высококвалифицированных стрелков тоническая активность мышц, обеспечивающих стояние, минимальна (на электромиограмме регистрируется наименьшая электрическая активность мышц). У спортсменов низкой квалификации и новичков наблюдается значительное напряжение этих мышц.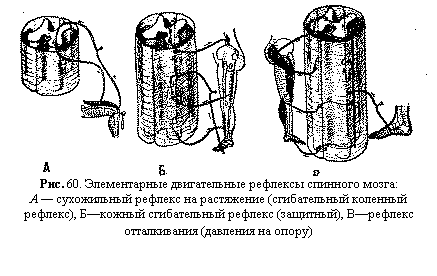 Миотатические и сухожильные рефлексы возникают при растяжении рецепторов мышц и сухожилий. Быстрое растягивание этих рецепторов рефлекторно вызывает фазное сокращение скелетных мышц. Это рефлекс активного противодействия мышцы ее растяжению. Рефлекторная дуга рефлекса на растяжение — моносинаптическая Она состоит всего из двух нейронов — афферентного и эфферентного, между которыми имеется лишь один синапс. Простота рефлекторной дуги обеспечивает быструю и точную реакцию мышцы, меньшую утомляемость рефлекторных ответов. Рефлексы на растяжение имеют большое значение при статической деятельности мышц для поддержания определенной позы, а также при осуществлении локомоторных актов (ходьбы, бега и др.). В произвольной двигательной деятельности человека иногда необходимо подавлять эти рефлексы (например, выполнение гимнастического шпагата требует подавления рефлекса на растяжение).
Миотатические и сухожильные рефлексы возникают при растяжении рецепторов мышц и сухожилий. Быстрое растягивание этих рецепторов рефлекторно вызывает фазное сокращение скелетных мышц. Это рефлекс активного противодействия мышцы ее растяжению. Рефлекторная дуга рефлекса на растяжение — моносинаптическая Она состоит всего из двух нейронов — афферентного и эфферентного, между которыми имеется лишь один синапс. Простота рефлекторной дуги обеспечивает быструю и точную реакцию мышцы, меньшую утомляемость рефлекторных ответов. Рефлексы на растяжение имеют большое значение при статической деятельности мышц для поддержания определенной позы, а также при осуществлении локомоторных актов (ходьбы, бега и др.). В произвольной двигательной деятельности человека иногда необходимо подавлять эти рефлексы (например, выполнение гимнастического шпагата требует подавления рефлекса на растяжение).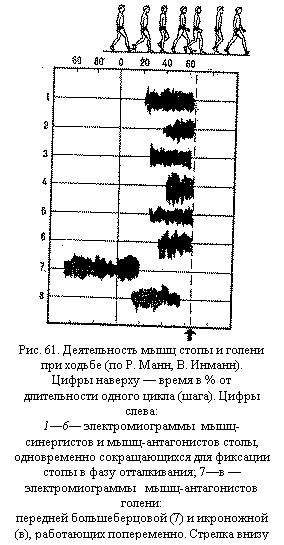 Значение коры больших полушарии в регуляции произвольных движений. Ведущую роль в регуляции произвольных движений играет кора больших полушарий, особенно ее высшие отделы—зоны перекрытия анализаторов, или третичные поля. В коре больших полушарий происходит выработка цели и задачи движений, соответственно этому строится и программа конкретных действий, которые нужны человеку для осуществления цели. В сложные поведенческие акты включаются не только моторные компоненты, но и необходимые вегетативные компоненты. Еще до начала движении кора больших полушарий повышает активность тех вставочных и моторных нейронов спинного мозга, которым предстоит участвовать в движении. В предстартовый период перед началом циклических движений в электрической активности коры происходит настройка на темп предстоящих движений. В тот момент, когда производится движение, кора тормозит деятельность всех посторонних афферентных путей и оказывается особенно восприимчивой к сигналам от рецепторов мышц, сухожилий и суставных сумок.
Значение коры больших полушарии в регуляции произвольных движений. Ведущую роль в регуляции произвольных движений играет кора больших полушарий, особенно ее высшие отделы—зоны перекрытия анализаторов, или третичные поля. В коре больших полушарий происходит выработка цели и задачи движений, соответственно этому строится и программа конкретных действий, которые нужны человеку для осуществления цели. В сложные поведенческие акты включаются не только моторные компоненты, но и необходимые вегетативные компоненты. Еще до начала движении кора больших полушарий повышает активность тех вставочных и моторных нейронов спинного мозга, которым предстоит участвовать в движении. В предстартовый период перед началом циклических движений в электрической активности коры происходит настройка на темп предстоящих движений. В тот момент, когда производится движение, кора тормозит деятельность всех посторонних афферентных путей и оказывается особенно восприимчивой к сигналам от рецепторов мышц, сухожилий и суставных сумок.


