
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Физиологические показатели функционального состояния нейронаСодержание книги
Поиск на нашем сайте Величина мембранного потенциала является основным параметром, который определяет значения важнейших показателей функционального состояния нейрона — его возбудимость и лабильность. Возбудимость нейрона. Возбудимость нейрона — это его способность отвечать на синоптическое воздействие потенциалом действия. Она зависит от соотношения двух параметров—мембранного потенциала и критического уровня деполяризации (рис. 46). В нормальных условиях деятельности критический уровень деполяризации нейрона относительно постоянный, поэтому возбудимость нейрона определяется в основном величиной мембранного потенциала. Подпороговые изменения мембранного потенциала. Мембранный потенциал может иметь нормальную величину (для нервной клетки—около 70 мв), уменьшенную (явление деполяризации) или большую (явление гиперполяризации). Для появления ПД при нормальном уровне мембранного потенциала требуется его сдвиг примерно на 10 мв, т. е. амплитуда порогового ВПСП должна быть около 10 мв для достижения критического уровня деполяризации. Эта величина и называется порогом возбудимости нейрона (см. рис. 46, Л). При активной деятельности нейрона в нем может некоторое время сохраняться состояние подпороговой деполяризации. Такое же состояние возникает при слабых синаптических воздействиях, когда появляются подподпороговые ВПСП. При этом разница между уровнем мембранного потенциала и критическим уровнем деполяризации уменьшается (см. рис. 46, Б), т. е. требуется уже меньшая сила раздражения (меньшая амплитуда порогового ВПСП) для появления ответного ПД. Возбудимость нейрона в этом случае повышена. При понижении возбудимости нервной клетки разница между уровнем мембранного потенциала и критическим уровнем деполяризации увеличена и, следовательно, требуется значительно большее, чем в норме, раздражение (большая амплитуда ВПСП) для появления ПД (см. рис. 46, В). Сверхпороговое изменение мембранного потенциала. При сильном возбуждении в нервной клетке могут возникать ВПСП очень большой амплитуды, которая значительно превосходит критический уровень деполяризации и длительное время сохраняется на этом сверхпороговом уровне. В результате в течение всего периода сверхпороговой деполяризации нейрон генерирует потенциалы действия (см. рис. 46, Г). Такие резкие сдвиги возбудимости нейронов возникают, например, под влиянием вышележащих отделов центральной нервной системы. Импульсы, приходящие к мотонейронам спинного мозга из головного мозга, способны вызывать мощные ВПСП с амплитудой 20—25 мв, которые гораздо выше пороговой величины (10 мв). При этом мотонейрон отвечает на одно раздражение залпом нескольких ПД. Этим достигается значительное усиление двигательного ответа при рефлекторных реакциях.
Изменения возбудимости нейрона при его деятельности. Протекание каждого ПД сопровождается закономерными изменениями возбудимости нейрона. В момент возникновения высоковольтной части — спайка нервная клетка не может ответить на раздражение новым ПД, т. е. является абсолютно невозбудимой. Эта фаза (абсолютная рефракторная фаза) длится примерно 0,5 мсек. Затем, в период постепенного снижения деполяризации мембраны, возбудимость нейрона восстанавливается до исходного уровня (фаза пониженной возбудимости, или относительная рефракторная фаза) и даже некоторое время может его превышать (супернормальная, или экзальтационная, фаза). После этого может наблюдаться еще некоторое понижение возбудимости во время развития следовой гиперполяризации (субнормальная фаза). Длительность описанных фаз изменения возбудимости определяет протяженность периодов повышенной активности (нормальная и супернормальная возбудимость) и пониженной активности (абсолютная и относительная рефракторные фазы, субнормальная фаза) нейронов. Так, большинство нейронов промежуточного мозга и коры больших полушарий после очередного разряда импульсов во время протекания абсолютной и относительной рефракторной фаз неактивны в течение примерно 100 мсек. Следовательно, наиболее удобным ритмом их спонтанной активности может быть ритм около 10 разрядов в 1 сек., который и является одним из факторов, определяющих частоту электрических колебаний на поверхности мозга в состоянии покоя. Лабильность нейрона. Лабильность нейрона—это скорость протекания элементарных реакций, лежащих в основе его возбуждения. Наиболее удобная мера лабильности нейрона—максимальная скорость протекания ПД. В качестве меры лабильности используют также максимальную частоту импульсов, воспроизводимых нейроном без трансформации в единицу времени, или оптимальную, наиболее устойчивую, частоту импульсов (текущая лабильность). Разные по функциям и величине нервные клетки обладают различными величинами лабильности. Даже в пределах одного нейрона лабильность разных его структур (дендритов, сомы, начального сегмента и аксона) резко различается. Величина лабильности нейрона определяется уровнем его мембранного потенциала. Лишь при определенной величине мембранного потенциала достигается оптимальный уровень возбудимости и лабильности нервной клетки, а также наиболее высокий уровень ее ритмической активности, что является важным условием для передачи информации нервной системе и осуществления целесообразных реакций. Например, наибольший максимальный ритм мотонейронов спинного мозга (до 100 импульсов в 1 сек.) отмечается при средней, оптимальной, величине мембранного потенциала—55 мв (т. е. при повышенной возбудимости мотонейрона). В естественных условиях деятельности частота разрядов мотонейрона обычно не превышает 50—60 импульсов в 1 сек. Лишь при мощных влияниях вышележащих отделов нервной системы резко изменяется мембранный потенциал мотонейронов и максимальная частота их разрядов может повыситься до 100 и даже до 300 импульсов в 1 сек. Однако разряды с такой частотой очень кратковременны. Они сменяются более медленным и устойчивым ритмом активности. В мелких промежуточных нейронах спинного мозга частота разрядов в начальные моменты активации достигает 600— 800 импульсов в 1 сек., а иногда превышает 1000 импульсов в 1 сек. Такая частота необходима для резкого и быстрого воздействия этих клеток на мотонейроны; через некоторое время она резко замедляется. Изменения в нейронах при активном и бездеятельном состояниях. Афферентные раздражения, возникающие при различных влияниях внешней и внутренней среды, в том числе при мышечной работе (с рецепторов мышц и сухожилий), повышают возбудимость и лабильность нейронов, их способность к стабильной ритмической активности. Бездеятельное же состояние снижает эти показатели, ухудшая функциональное состояние нервной клетки. Активная деятельность клетки вызывает не только функциональные изменения, но и значительные структурные и биохимические перестройки различных ее элементов. Прогрессивные изменения наблюдаются в синаптическом аппарате нейронов. Эксперименты с тренировкой животных (тренировка задних конечностей собаки путем массажа, электрического раздражения и пассивной гимнастики, тренировка крыс с помощью плавания) показали, что средняя величина (диаметр) синапсов на мотонейронах поясничного отдела спинного мозга достоверно возрастает (примерно на 35%). С увеличением размеров синапсов улучшается проведение нервных импульсов по рефлекторным путям. Активное участие корковых нейронов в условнорефлекторной деятельности вызывает усиленное развитие шипикового аппарата на дендритах. Это способствует улучшению межнейронных взаимосвязей в коре больших полушарий. Подавление условнорефлекторной деятельности различными фармакологическими воздействиями приводит к ухудшению рецепторной функции нейронов — нарушению синаптических контактов (исчезновению шипиков), а затем и к Деформации дендритов. При усилении афферентной импульсации, поступающей к нейрону, в нейрональных синапсах происходят увеличение числа синаптических пузырьков, их усиленное новообразование и интенсивное перемещение в оперативную зону пресинаптической мембраны. Кроме того, в пресинаптических образованиях отмечается еще большее усиление (в 7 раз по сравнению с покоем) синтеза медиатора (ацетилхолина) и выраженное увеличение его выделения (в 70 раз). В нейронах повышается уровень окислительных процессов, возрастает количество белков и РНК в телах нервных клеток и в клетках глии. При усиленной двигательной активности увеличиваются размеры эффекторных нейронов—пирамидных клеток коры и мотонейронов спинного мозга. Все прогрессивные структурные, биохимические и функциональные изменения улучшают осуществление нервными клетками их сложных функций. Это лежит в основе совершенствования различных поведенческих реакций целого организма, в том числе двигательной деятельности и психических процессов у человека. Системы нервных клеток Диффузная нервная сеть и процесс централизации. У простейших одноклеточных животных одна клетка осуществляет разнообразные функции. Передвижение этих животных обеспечивается либо выпячиванием части клетки (ложноножки) и переливанием в нее содержимого клетки (амебоидное движение, характерное и для белых клеток крови человека), либо с помощью специальных образований — ресничек или жгутиков. Усложнение деятельности организма привело к разделению функций различных клеток — их специализации. Одни из них приобрели способность к сокращению (мышечные клетки), другие — к восприятию внешних и внутренних раздражении, переработке поступающей информации и передаче управляющих сигналов на органы движения и другие органы тела (нервные клетки). Наиболее простая форма организации нервной системы — диффузная нервная сеть низших беспозвоночных животных (губок, гидр, актиний и медуз). нейроны такой сети имеют многочисленные взаимные связи, по которым возбуждение распространяется диффузно, по всем направлениям. Более сложной формой является сегментарная организация нервной системы у высших беспозвоночных животных (червей, насекомых). Тело их состоит из имеющих одинаковое строение участков — сегментов, иннервация которых осуществляется расположенными в этих же сегментах нервными клетками. С усложнением поведенческих реакций происходит развитие у позвоночных животных головного конца нервной системы—энцефалона. В нем сосредоточиваются группы нервных клеток, управляющих важнейшими функциями, — нервные центры. При этом утрачивается автономность отдельных сегментов и все большая часть функций передается вышележащим отделам нервной системы. Этот процесс получил название энцефализации (централизация) функций. С формированием коры больших полушарий происходит подчинениевсех других отделов нервной системы, т. е. процесс кортикализации. Строение и функции центральной нервной системы у человека отражают этот эволюционный процесс. Более древние и менее сложно организованные ее отделы имеют более выраженную, сегментарную организацию. Это спинной мозг, продолговатый и средний, многие центры которых иннервируют расположенные на том же уровне участки тела (рис. 47). Находящиеся над ними более мо Соматотопическое представительство функций. Системы нейронов, образующих восходящие пути от рецепторов или нисходящие "ути к рабочим органам, расположены в определенном порядке — по типу проекции «точку в точку». Так, каждый участок воспринимающей поверхности глаза (сетчатой оболочки с фоторецепторами) передает свои сигналы определенному участку в зрительной области коры больших полушарий. Эти чувствительные корковые центры расположены таким образом, что они как бы образуют экран, отражающий расположение фоторецепторов на периферии. Следовательно, наш мозг получает объективную информацию о пространственных особенностях внешнего мира. Определенная пространственная организация в нервной системе сигналов от различных органов чувств (от зрительных, слуховых, мышечных, тактильных и других рецепторов) позволяет человеку иметь необходимое представление о схеме пространства. Таким же образом в мозгу обеспечивается представление о схеме тела. Порядок размещения афферентных волокон в проводящих путях и локализация нервных центров в проекционных областях разных этажей нервной системы соответствуют порядку размещения участков кожи и скелетных мышц в теле. Этому принципу отвечает представительство моторных функций в различных этажах нервной системы. Так, например, в самой верхней части передней центральной извилины коры больших полушарий находятся центры, посылающие импульсы к мышцам нижних конечностей, несколько ниже — моторные центры мышц туловища, еще ниже—моторное представительство мышц верхних конечностей и, наконец, центры мышц шеи и головы. Таков же порядок расположения этих центров в моторных ядрах нижележащих этажей нервной системы (например, в красном ядре среднего мозга). Такая организация связей в центральной нервной системе, при которой пространственное расположение нейронов, связанных с определенными рецепторами или рабочими органами на периферии, отражает специфику их пространственного расположения в организме, называется соматопическим (от лат. somaticus — телесный и topicus — местный) или экранным представительством функций. В соматопической организации нервной системы важное значение имеют: 1) функциональная значимость отдельного органа тела и 2) роль данного участка мозга в его иннервации. В связи с этим возникает несоответствие размеров органа тела и размеров территории мозга, которая управляет его деятельностью. Например, мышцы пальцев руки выполняют чрезвычайно сложные функции в произвольных движениях человека. В их деятельности основную роль играют импульсы, идущие от коры больших полушарий. Этим объясняется то, что представительство (как сенсорное, так и в особенности моторное) этих мышц в коре занимает значительно большую территорию, чем представительство всех остальных мышц тела. Механизмы взаимодействия нервных клеток. Нервные клетки функционируют в тесном взаимодействии друг с другом. Значение нервных импульсов. Все взаимодействия между нервными клетками осуществляются благодаря двум механизмам: 1) влияниям электрических полей нервных клеток (электротоническим влияниям) и 2) влияниям нервных импульсов. Первые распространяются на очень небольшие территории мозга Электрический заряд нервной клетки создает вокруг нее электрическое поле, колебания которого вызывают изменения электрических полей лежащих рядом нейронов, что приводит к изменениям их возбудимости, лабильности и проводимости. Электрическое поле нейрона имеет сравнительно небольшую протяженность—около 100 мк, оно быстро затухает по мере удаления от клетки и может оказывать воздействие лишь на соседние нейроны. Второй механизм обеспечивает не только ближайшие взаимодействия, но и передачу нервных влияний на большие расстояния. Именно с помощью нервных импульсов происходит объединение отдаленных и изолированных участков мозга в общую, синхронно работающую систему, что необходимо для протекания сложных форм деятельности организма. Нервный импульс, следовательно, является основным средством связи между нейронами. Высокая скорость распространения импульсов и локальное их воздействие на избранную точку мозга способствуют быстрой и точной передаче информации в нервной системе. В межнейронных взаимодействиях используется частотный код, т. е. изменения функционального состояния и характера ответных реакций одной нервной клетки кодируются изменением частоты импульсов (потенциалов действия), которые она посылает к другой нервной клетке. Общее количество импульсов, отправляемых нервной клеткой в единицу времени, или ее суммарная импульсная активность,—важный физиологический показатель деятельности нейрона. Значение синапсов. В передаче нервных импульсов от одного нейрона к другому большое значение имеют синапсы. Функционально возбуждение передается только через синапсы. Они выступают как переключатели, которые могут быть еще не включены (до образования долговременной проводимости-памяти) или уже хорошо проводить возбуждение, запуская от пришедшего стимула запомненную связь с ним последующих звеньев, что и обуславливает механизм долговременной памяти. Особенно много их в высших отделах нервной системы и у нейронов с наиболее сложными функциями. С совершенствованием условнорефлекторной деятельности в процессе эволюции животных организмов увеличивается число синаптических контактов на пирамидных нейронах коры больших полушарий, в частности за счет увеличения числа шипиков на их дендритах. Состояние синапсов на этих шипиках коррелирует с состоянием высшей нервной деятельности. При подавлении условных рефлексов некоторыми веществами (например, аминазином) контакты на шипиках разрушаются, исчезают сами шипики, а затем деформируются и дендритные отростки нейрона. Способность синапсов передавать нервные импульсы непостоянна. Она повышается после активной деятельности синапса и снижается при отсутствии активности. Понижение функциональных возможностей синапсов (гипосинапсия) ведет к ухудшению проведения через них нервных импульсов, а их полное нарушение (асинапсия) вызывает окончательное разобщение нервных клеток. При отсутствии активации синапса в течение нескольких дней уже наблюдаются уменьшение размера синаптических пузырьков, запасов медиатора, нарастание длительности синаптической задержки и т. п. Бездействие синаптических контактов на мотонейронах спинного мозга, вызванное перерезкой чувствительных путей от соответствующих мышечных волокон, через 2—4 недели уменьшает их возбуждающее влияние на мембрану мотонейрона вдвое по сравнению с влияниями других афферентных путей, оканчивающихся на том же мотонейроне. У кролика через несколько дней содержания в абсолютной темноте, т. е. после полного бездействия рецепторных клеток глаза, уменьшаются синаптические пузырьки в окончаниях их аксонов. Это объясняет постепенное угасание приобретенных навыков (хотя механизм забывания этим не исчерпывается). Длительность этого процессам может быть от нескольких минут до десятков лет, в зависимости от локализации (см ниже). Активная деятельность нервных клеток способствует улучшению синоптического взаимодействия между ними: росту площади синаптических контактов, ускорению проведения через синапсы и т. д. Это наблюдается, например, при двигательной деятельности. Как показали эксперименты на животных, ежедневная принудительная ходьба их по 1 часу приводит к значительной активации синапсов в соответствующих рефлекторных дугах спинного мозга. Синапсы, являются самым чувствительным местом нервной системы. Именно синаптические контакты между нейронами первыми реагируют на различные повреждающие воздействия. При травмах нервной ткани, отеках, нарушениях кровообращения, рентгеновских облучениях, отравлениях и других неблагоприятных воздействиях в синапсах сначала возникают обратимые изменения — сильное набухание (увеличение их размеров в несколько раз) и деформация, а затем происходят необратимые изменения синапсов — их разрушение, в результате чего полностью нарушается нервная деятельность. Свойства нервных центров Понятие о нервном центре. В сложных многоклеточных организмах животных и человека отдельная нервная клетка не в состоянии регулировать какие-либо функции. Все основные формы деятельности нервной системы связаны с участием в ее функциях определенных групп нервных клеток—нервных центров. Нервным центром называют совокупность нервных клеток, необходимых для осуществления какой-либо функции. Эти центры отвечают соответствующими рефлекторными реакциями на внешнее раздражение, поступившее от связанных с ними рецепторов. Клетки нервных центров реагируют и на непосредственное их раздражение веществами, находящимися в протекающей через них крови (гуморальные влияния). Для выявления функций нервного центра используют ряд методов: метод раздражения (усиление его деятельности), метод разрушения или удаления — экстирпации (нарушение регулируемой им функции), электрофизиологический метод — регистрация электрических явлений в нервном центре и др. Так, функциональное значение, а также локализация дыхательного центра в продолговатом мозгу были выявлены путем последовательных перерезок головного мозга. При перерезках выше области его расположения (выше продолговатого мозга) дыхание сохранялось, а при перерезках ниже продолговатого мозга дыхательные движения исчезали. Сложные реакции в целостном организме обычно связаны с участием многих нервных центров, расположенных в различных этажах центральной нервной системы. Например, произвольное изменение дыхания осуществляется человеком при участии центров коры больших полушарий, особого центра в среднем мозгу, дыхательного центра продолговатого мозга и центров спинного мозга, иннервирующих дыхательные мышцы. Физиологические свойства нервных центров определяют характер ответных реакций. Эти свойства в значительной мере связаны с особенностями проведения нервных импульсов через синапсы, соединяющие различные нервные клетки. Одностороннее проведение возбуждения. В отличие от нервного волокна, в котором возможно двустороннее проведение волны возбуждения от места нанесения раздражения, в нервных центрах волна возбуждения проводится только в одном направлении: раздражение задних (чувствительных) корешков спинного мозга вызывает возникновение нервного импульса в передних двигательных корешках, однако при раздражении передних корешков волна возбуждения в задних корешках не появляется. Это обусловлено особенностями синаптической передачи возбуждения в отдельных нервных клетках. Поскольку проведение волны возбуждения от одного нейрона к другому через синапс происходит химическим путем—с помощью медиатора, а медиатор содержится лишь в пресинаптической части синапса и отсутствует в постсинаптической мембране, — проведение нервных влияний через синапс возможно только от пресинаптической мембраны к постсинаптической и невозможно в обратном направлении (см. рис. 45). В связи с этим поток нервных импульсов в рефлекторной дуге имеет определенное направление: от афферентных нейронов к вставочным и затем к эффекторным — мотонейронам и вегетативным нейронам. Замедленное проведение возбуждения. В основе проведения нервных импульсов по цепочке нейронов лежат два различных механизма: электрический (проведение потенциала действия — ПД — по нервным волокнам) и химический (передача через синапс с помощью медиатора). Первый осуществляется с большой скоростью (до 100—140 м/сек), второй—в тысячу раз медленнее. Замедление проведения связано с затратой времени на процессы, происходящие от момента прихода пресинаптического импульса в синапс до появления в постсинаптической мембране возбуждающих или тормозных потенциалов. Этот интервал называется синаптической задержкой и составляет в мотонейронах и большинстве других клеток Центральной нервной системы примерно 0,3 мсек. В вегетативной нервной системе длительность синаптической задержки больше — не менее 10 мсек. За это время пресинаптический импульс вызывает опорожнение синаптических пузырьков, происходит диффузия медиатора через синаптическую щель, увеличение под его влиянием ионной проницаемости постсинаптической мембраны и возникает постсинаптический потенциал. После этого требуется еще около 1,2 мсек. на развитие возбуждающего постсинаптического потенциала (ВПСП) до максимума, чтобы возник (в случае достижения критического уровня деполяризации) ПД. Таким образом, весь процесс передачи нервного импульса (от ПД одной клетки до ПД следующей клетки) через один синапс занимает примерно 1,5 мсек. При утомлении, охлаждении и ряде других воздействий длительность синаптической задержки возрастает. Если же для осуществления какой-либо реакции требуется участие большого числа нейронов (многих сотен и даже тысяч), то суммарная величина задержки проведения по нервным центрам — так называемое центральное время проведения — может составить десятые доли секунды и даже целые секунды. Поскольку проведение импульсов по нервным
Суммация. В ответ на одиночную афферентную волну, идущую от мышц к мотонейронам, в пресинаптической части синапса освобождается 1 квант нейромедиатора. При этом в пресинаптической мембране мотонейрона обычно возникает ВПСП подпороговой величины – 0,1 – 0,2 мв, недостаточной для вызова ответной реакции клетки. Чтобы величина ВПСП достигла критического уровня (порога возникновения ПД – примерно 10 мв.), требуется суммация на мембране клетки многих подпороговых ВВСП. Различают пространственную и временную суммацию. Пространственная суммация наблюдается в случае одновременного поступления нескольких импульсов в один и тот же нейрон по разным пресинаптическим волокнам. Одновременное возбуждение синапсов в различных участках мембраны мотонейрона повышает амплитуду суммарного ВПСП до пороговой величины, возникает ответный импульс мотонейрона и рефлекторно осуществляется двигательная реакция. Так, при синхронном раздражении трех афферентных нервах волокон—от короткой малоберцовой мышцы, длинного разгибателя пальцев и длинной малоберцовой мышцы — в мотонейроне длинной малоберцовой мышцы регистрируется ВПСП, равный сумме отдельных ВПСП, получаемых при изолированном раздражении каждого волокна (рис. 49, Б}. Обычно требуется одновременная активация 50—100 афферентных волокон для получения ответа двигательной клетки спинного мозга. Временная суммация происходит при активации одного и того же афферентного пути серией последовательных раздражении. Если интервалы между поступающими импульсами достаточно коротки и ВПСП мотонейрона от предыдущего раздражения не успевают затухать, то последующие ВПСП накладываются друг на друга, пока деполяризация мембраны не Достигнет критического уровня для возникновения ПД (см. рис. 49, Л). Явления суммации могут наблюдаться при осуществлении отдельных рефлексов (например, рефлексов чихания и кашля в ответ на слабые раздражения слизистой оболочки дыхательных путей). Они лежат в основе формирования нейронных комплексов, определяющих поведение целого организма, в основе развития условных рефлексов. Фоновая активность. Многие нервные клетки обладают способностью к постоянной импульсной активности. Их мембранный потенциал периодически колеблется, то увеличиваясь, то уменьшаясь. Каждый раз, когда его увеличение превышает критический уровень деполяризации (порог возбудимости клетки), возникает импульсный разряд. Такая автоматическая деятельность называется фоновой активностью нейрона (рис. 50). Ее могут вызывать случайные импульсы, циркулирующие в нервной системе, но она может проявляться и при отсутствии приходящих к нейрону раздражении—в результате изменении ионных и обменных процессов, колебаний температуры и других причин. В последнем случае ее называют спонтанной.
бездеятельные. Это облегчает протекание многих рефлекторных реакций. Во-вторых, клетки, обладающие постоянным ритмом фоновой активности, могут иметь значение водителя ритма (пейсмекера), т. е. задавать этот ритм другим нервным клеткам. При этом возникают большие комплексы синхронно работающих клеток, в которых благодаря общему ритму особенно хорошо выражено взаимодействие между отдельными нейронами. В-третьих, некоторые такие клетки выполняют роль счетчиков времени в центральной нервной системе. Их деятельность помогает поддерживать определенный ритм движений, субъективно оценивать интервалы времени, через определенные сроки включать необходимые функции и т. п. Автоматическая деятельность присуща клеткам дыхательного центра. Их спонтанные разряды позволяют поддерживать ритмические дыхательные движения не только при бодрствовании, но и во время сна человека, в условиях наркоза; периодические разряды дыхательного центра регистрируются даже при полной изоляции продолговатого мозга от других отделов центральной нервной системы.
Усвоение и трансформация ритма. При ритмических раздражениях вызванная активность нейрона может настроиться на ритм приходящих импульсов (явление усвоения ритма—рис. 51). Эта настройка деятельности нейрона на заданный извне ритм (реакция усвоения, перестройки или навязывания ритма) имеет большое значение для организации взаимодействия между различными нервными центрами и различными этажами нервной системы, в частности для организации ритмических движений у человека. Активность нейрона может перестроиться как на более высокий, так и на более низкий ритм. В результате такой способности многие нервные клетки получают возможность сонастраиваться, т. е. работать сообща, в одном комплексе, не мешая друг другу различной по темпу импульсацией. Впервые вопрос о значении усвоения ритма для протекания реакций организма был поставлен А. А. Ухтомским и его учениками (Н. В. Голиковым, Е. К. Жуковым и др.). Особенно важно, что в процессе ритмического раздражения рефлекторной дуги нервные центры способны усвоить ритм активности, превышающий их первоначальную лабильность. Это явление лежит в основе повышения работоспособности отдельных рефлекторных дуг, а также целостного организма (в частности, это имеет место в спортивной тренировке). Очень часто вызванная активность нейрона не соответствует ритму приходящей к данной клетке импульсации. Нервные клетки обладают свойством изменять частоту передающихся импульсов, т.е. свойством трансформации ритма. Характер ответного разряда зависит, во-первых, от свойств раздражителя и, во-вторых, от функционального состояния нейрона (его мембранного заряда, возбудимости, лабильности). В обычных условиях чем сильнее раздражение, тем чаще проявляется ответная импульсация. При высокой возбудимости нейрона может возникать учащение импульсации (мультипликация ритма). В этом случае афферентная волна вызывает длительные надпороговые ВПСП в клетке, и на.каждое одиночное раздражение клетка отвечает залпом из нескольких ПД (пока ВПСП превышает пороговый уровень). При низком уровне возбудимости и лабильности нейрона чаще происходит урежение ритма, так как несколько приходящих импульсов должны суммироваться, чтобы наконец достичь порога возникновения ПД. В результате на несколько приходящих импульсов нейрон отвечает лишь одним импульсом. В некоторых крупных клетках, например мотонейронах спинного мозга, имеются специальные механизмы, регулирующие ритм ответных реакций. После каждого ПД в этих клетках наблюдаются выраженные следовые процессы — следовая гиперполяризация мембраны. В этот момент
|
||
|
|
Последнее изменение этой страницы: 2016-09-20; просмотров: 1030; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.108 (0.013 с.) |

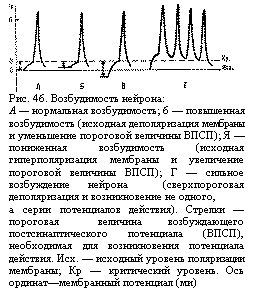 нейрона: А — нормальная возбудимость; 6 — повышенная возбудимость (исходная деполяризация мембраны и уменьшение пороговой величины ВПСП); Я — пониженная возбудимость (исходная гиперполяри-зация мембраны и увеличение пороговой величи-ны ВПСП); Г — сильное возбуждение нейрона (сверхпороговая деполяризация и возникновение не одного, а серии потенциалов действия). Стрелки — пороговая величина возбуждающего постсинаптического потенциала (ВПСП), необходимая для возникновения потенциала действия. Исх. — исходный уровень поляризации мембраны; Кр — критический уровень. Ось ординат—мембранный потенциал (ми) ">Величина деполяризации нервных клеток находится в линейной зависимости от частоты, раздражающих импульсов. Высшие отделы мозга, посылая импульсы различной частоты к нейронам нижележащих отделов, регулируют их возбудимость, осуществляя контроль за ответными реакциями организма.
нейрона: А — нормальная возбудимость; 6 — повышенная возбудимость (исходная деполяризация мембраны и уменьшение пороговой величины ВПСП); Я — пониженная возбудимость (исходная гиперполяри-зация мембраны и увеличение пороговой величи-ны ВПСП); Г — сильное возбуждение нейрона (сверхпороговая деполяризация и возникновение не одного, а серии потенциалов действия). Стрелки — пороговая величина возбуждающего постсинаптического потенциала (ВПСП), необходимая для возникновения потенциала действия. Исх. — исходный уровень поляризации мембраны; Кр — критический уровень. Ось ординат—мембранный потенциал (ми) ">Величина деполяризации нервных клеток находится в линейной зависимости от частоты, раздражающих импульсов. Высшие отделы мозга, посылая импульсы различной частоты к нейронам нижележащих отделов, регулируют их возбудимость, осуществляя контроль за ответными реакциями организма.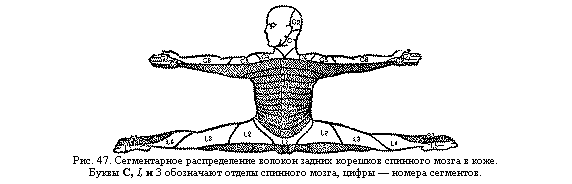 лодые отделы, так называемые надсегментарные—промежуточный мозг, мозжечок, кора больших полушарий,—не имеют непосредственной связи с органами тела, а управляют их деятельностью через сегментарные центры. Надсегментарные отделы приобретают по мере совершенствования все более важную роль в жизнедеятельности органов. Они не только подчиняют, но и значительно перестраивают функции нижележащих структур, оставляя им лишь самые элементарные рефлексы. Так, отделение спинного мозга от головного вызывает угнетение всех его рефлексов—спинальный шок, который выражен гораздо слабее и длится меньше (всего несколько минут) У рыб и земноводных и более глубок и длителен (несколько недель) у млекопитающих. После исчезновения явлений шока спинальная лягушка (лишенная головного мозга) может осуществлять довольно сложные координированные движения (например, удаление пальцами задней лапки кислотного раздражителя с бедра), у спинальных кошек и собак могут наблюдаться сгибание, разгибание, маятникообразные движения конечностей, а у человека при перерыве связи головного мозга со спинным — практически только сгибательные рефлексы (разгибательные движения у него ниже места перерыва спинного мозга почти не наблюдаются, перекрестный разгибательный рефлекс выражен намного слабее, чем у спинальной собаки).
лодые отделы, так называемые надсегментарные—промежуточный мозг, мозжечок, кора больших полушарий,—не имеют непосредственной связи с органами тела, а управляют их деятельностью через сегментарные центры. Надсегментарные отделы приобретают по мере совершенствования все более важную роль в жизнедеятельности органов. Они не только подчиняют, но и значительно перестраивают функции нижележащих структур, оставляя им лишь самые элементарные рефлексы. Так, отделение спинного мозга от головного вызывает угнетение всех его рефлексов—спинальный шок, который выражен гораздо слабее и длится меньше (всего несколько минут) У рыб и земноводных и более глубок и длителен (несколько недель) у млекопитающих. После исчезновения явлений шока спинальная лягушка (лишенная головного мозга) может осуществлять довольно сложные координированные движения (например, удаление пальцами задней лапки кислотного раздражителя с бедра), у спинальных кошек и собак могут наблюдаться сгибание, разгибание, маятникообразные движения конечностей, а у человека при перерыве связи головного мозга со спинным — практически только сгибательные рефлексы (разгибательные движения у него ниже места перерыва спинного мозга почти не наблюдаются, перекрестный разгибательный рефлекс выражен намного слабее, чем у спинальной собаки). волокнам от периферических рецепторов в нервные центры и от нервных центров к исполнительным органам занимает сравнительно небольшое время, общее время от момента нанесения внешнего раздражения до появления ответной реакции организма (латентный период рефлекса) определяется центральным временем проведения. Величина латентного периода рефлекса служит важным показателем функционального состояния нервных центров и широко используется в практике. Латентный период двигательной реакции измеряется от момента подачи сигнала до момента начала движения (или до появления электрической активности соответствующей мышцы). При осуществлении человеком простых движений в ответ на внешние сигналы, например нажимание на кнопку при световом сигнале (рис. 48, А), длительность латентного периода двигательной реакции составляет около 120 – 220 мсек., а при утомлении, недостатке кислорода и пр. она может увеличиваться до 300 – 500 и более миллисекунд. При более сложных движениях, например выполнении фехтовальщиком укола в мишень с выпадом в ответ на световой сигнал, этот период значительно больше – около 300 – 400 мсек. и более (см. рис. 48, Б).
волокнам от периферических рецепторов в нервные центры и от нервных центров к исполнительным органам занимает сравнительно небольшое время, общее время от момента нанесения внешнего раздражения до появления ответной реакции организма (латентный период рефлекса) определяется центральным временем проведения. Величина латентного периода рефлекса служит важным показателем функционального состояния нервных центров и широко используется в практике. Латентный период двигательной реакции измеряется от момента подачи сигнала до момента начала движения (или до появления электрической активности соответствующей мышцы). При осуществлении человеком простых движений в ответ на внешние сигналы, например нажимание на кнопку при световом сигнале (рис. 48, А), длительность латентного периода двигательной реакции составляет около 120 – 220 мсек., а при утомлении, недостатке кислорода и пр. она может увеличиваться до 300 – 500 и более миллисекунд. При более сложных движениях, например выполнении фехтовальщиком укола в мишень с выпадом в ответ на световой сигнал, этот период значительно больше – около 300 – 400 мсек. и более (см. рис. 48, Б). Укорочение латентного периода одной и той же двигательной реакции человека в процессе спортивной тренировки может отражать повышение функционального состояния нервных центров (ускорение проведения) и перестройку нервных цепей, участвующих в передаче нервных импульсов (укорочение проводящего пути).
Укорочение латентного периода одной и той же двигательной реакции человека в процессе спортивной тренировки может отражать повышение функционального состояния нервных центров (ускорение проведения) и перестройку нервных цепей, участвующих в передаче нервных импульсов (укорочение проводящего пути). Фоновая активность играет важную роль в деятельности целого организма. Во-первых, импульсы, приходящие по афферентным путям, легче возбуждают клетки со спонтанной активностью, чем
Фоновая активность играет важную роль в деятельности целого организма. Во-первых, импульсы, приходящие по афферентным путям, легче возбуждают клетки со спонтанной активностью, чем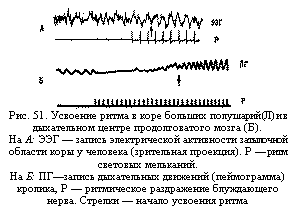 Вызванная активность. Импульсные разряды нейрона, возникающие в ответ на внешнее раздражение, называются вызванной активностью. Ответ нервной клетки может возникнуть в форме одиночного ПД, серии импульсов с затухающей частотой, а также в виде пачек импульсов, появляющихся через определенные интервалы. В различных нейронах наблюдаются разные типы вызванной активности. Одни клетки дают разряды только в момент включения раздражителя (эффект включения) —это наиболее частая реакция клеток, другие — только в момент его выключения (эффект выключения), третьи—в обоих случаях. В центральной нервной системе имеются клетки, дающие разряды лишь на новые раздражители, что обеспечивает ориентировочную реакцию. Известны также клетки, в которых отмечается вызванная активность при неправильных реакциях организма, т. е. сигнализирующие об ошибках, что позволяет вносить коррекции в действия человека.
Вызванная активность. Импульсные разряды нейрона, возникающие в ответ на внешнее раздражение, называются вызванной активностью. Ответ нервной клетки может возникнуть в форме одиночного ПД, серии импульсов с затухающей частотой, а также в виде пачек импульсов, появляющихся через определенные интервалы. В различных нейронах наблюдаются разные типы вызванной активности. Одни клетки дают разряды только в момент включения раздражителя (эффект включения) —это наиболее частая реакция клеток, другие — только в момент его выключения (эффект выключения), третьи—в обоих случаях. В центральной нервной системе имеются клетки, дающие разряды лишь на новые раздражители, что обеспечивает ориентировочную реакцию. Известны также клетки, в которых отмечается вызванная активность при неправильных реакциях организма, т. е. сигнализирующие об ошибках, что позволяет вносить коррекции в действия человека.


