Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Основные механизмы регуляции дыханияСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
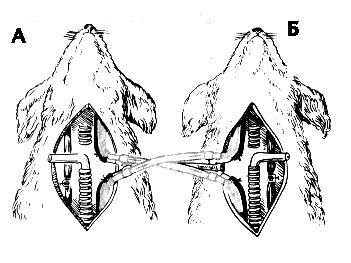
Интенсивность тканевого дыхания изменяется в широких пределах в соответствии с деятельностью органов и тканей, следовательно, изменяется и потребность в снабжении кислородом, и количество выделяемого углекислого газа. Адекватное снабжение клеток кислородом и удаление углекислого газа из организма осуществляется координированной деятельностью всех перечисленных систем, изменения происходят на всех этапах транспорта газов, приспосабливая темп окислительных процессов к меняющимся условиям внешней среды. Некоторые изменения происходят очень быстро, например способность гемоглобина присоединять или отдавать кислород в соответствии с его напряжением в крови, активность окислительных ферментов, быстро происходит рефлекторное изменение минутного объема дыхания, деятельности сердца, тонуса сосудов - эти изменения происходят в течение секунд. Больше времени требуется для изменения, например, количества эритроцитов в крови за счет выброса их из депо, объема циркулирующей крови. Еще более длительно протекают процессы структурной перестройки, требующие синтеза белка, такие как эритропоэз синтез гемоглобина, увеличение числа митохондрий в клетках. Таким образом, регуляция дыхания осуществляется на нескольких уровнях, которые включаются не одновременно, и вносят не одинаковый вклад в конечный результат, однако нарушение любого из этих процессов может лежать в основе патологических процессов дыхательной системы. Учитывая то, что вклад отдельных компонентов системы в процесс доставки кислорода тканям неодинаков, попытаемся выделить главный механизм. Диффузия газов в ткани и из тканей осуществляется пассивно, в соответствии с градиентом напряжения газов между капиллярами и тканевой жидкостью, следовательно, зависит от интенсивности метаболизма в тканях, объемной и линейной скорости тока крови, концентрации гемоглобина, кислотно-щелочного равновесия. Однако, как бы ни были значимы эти параметры, если кровь не будет достаточно насыщена кислородом, или будет содержать избыток углекислоты, диффузия окажется невозможной. Следовательно, для нормального протекания тканевого обмена особенно важны содержание О2 и СО2 в артериальной крови. Известно, что при протекании крови через капилляры легких между плазмой и альвеолярным воздухом устанавливается почти полное газовое равновесие, поэтому состав альвеолярного воздуха определяет содержание О2 и СО2 в артериальной крови. Оптимальное содержание газов в альвеолярном воздухе поддерживается изменением объема легочной вентиляции в соответствии с ситуацией в организме в определенный момент. Таким образом, на внешнее дыхание ложится задача обеспечить оптимальные величины О2 и СО2 в крови при любом уровне тканевого метаболизма. Регуляция внешнего дыхания Вентиляция легких - это процесс обновления газового состава альвеолярного воздуха, который обеспечивает поступление кислорода и выведение углекислого газа. Этот процесс осуществляется ритмичной работой дыхательных мышц, изменяющих объем грудной клетки. Интенсивность вентиляции определяется глубиной вдоха и частотой дыхания. Таким образом, минутный объем дыхания - это показатель легочной вентиляции, который должен обеспечивать тот газовый гомеостаз, который необходим в конкретной ситуации (покой, физическая работа). Регуляция внешнего дыхания представляет собой процесс изменения минутного объема дыхания в различных условиях для обеспечения оптимального газового состава внутренней среды организма. Во второй половине ХIX века появилась гипотеза о том, что основными факторами регуляции дыхания являются парциальное давление кислорода и углекислого газа в альвеолярном воздухе и, следовательно в артериальной крови. Экспериментальное доказательство того, что обогащение артериальной крови углекислотой и обеднение кислородом усиливает вентиляцию легких в результате наступающего при этом возбуждения дыхательного центра, было получено в классическом опыте Фредерика с перекрестным кровообращением в 1890г (рисунок 13). У двух собак, находившихся под наркозом, перерезали и соединяли перекрестно сонные артерии и отдельно яремные вены. После такого соединения и перевязки позвоночных артерий голова первой собаки снабжалась кровью второй и наоборот. Если у первой собаки перекрывали трахею и вызывали таким путем асфиксию, то у второй собаки развивалось гиперпноэ - увеличение легочной вентиляции. У первой же собаки, несмотря на увеличение в крови напряжения углекислоты и снижение напряжения кислорода через некоторое время наступало апноэ - прекращение дыхательных движений. Это объясняется тем, что в сонную артерию первой собаки поступает кровь второй собаки, у которой в результате гипервентиляции снижается содержание углекислоты в артериальной крови. Уже тогда было установлено, что регуляция дыхания происходит путем обратной связи: отклонения в газовом составе артериальной крови приводят путем воздействия на дыхательный центр такие изменения дыхания, которые уменьшают эти отклонения.
Рисунок 13. Схема опыта Фредерика с перекрестным кровообращением Пережатие трахеи у собаки А вызывает одышку у собаки Б. Одышка у собаки Б вызывает замедление и остановку дыхания у собаки А
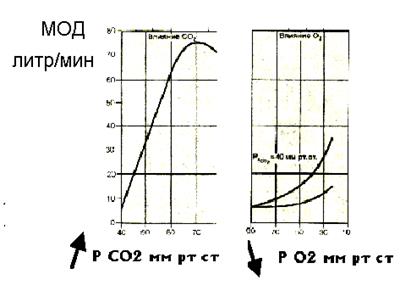
Дальнейшее развитие «гуморальной» теории дыхания (теории управления по отклонению напряжения газов в крови) связано с именем Холдейна. Он разработал методы получения и анализа альвеолярного воздуха у человека и установил, что парциальные давления газов в альвеолярном воздухе обычно колеблются в очень узких пределах. Холдейн пришел к выводу, что основным фактором регуляции дыхания является напряжение углекислоты в артериальной крови. Работы Холдейна получили широкое признание, а его главный вывод о том, что повышение напряжения углекислоты в артериальной крови приводит к большому увеличению МОД, остался справедливым вплоть до настоящего времени. Важная роль углекислоты в регуляции дыхания может быть проиллюстрирована тем, что увеличение содержания СО 2 в альвеолах на 0,2 % вызывает увеличение вентиляции легких на 100 %. Еще в начале ХIX века было показано, что в продолговатом мозге на дне IV желудочка расположены структуры, разрушение которых уколом иглы ведет к прекращению дыхания и гибели организма. Этот небольшой участок мозга в нижнем углу ромбовидной ямки был назван дыхательным центром. Многочисленными исследованиями удалось установить, что изменения газового состава внутренней среды оказывают влияние на дыхательный центр не непосредственно, а путем воздействия на специальные хемочувствительные рецепторы, расположенные в продолговатом мозге - центральные (медуллярные) хеморецепторы и в сосудистых рефлексогенных зонах - периферические (артериальные) хеморецепторы. Хотя основной целью дыхания является доставка клеткам кислорода, вентиляция легких управляется преимущественно в соответствии с продукцией в организме двуокиси углерода, т.е. поддерживает в основном не кислородный, а углекислотный (или, что почти одинаково, водородный) гомеостаз. Такие принципы сформировались в ходе заселения высшими позвоночными суши и перехода от водного дыхания к воздушному. Вспомним, что у рыб важнейшим фактором, регулирующим дыхательные движения, служит напряжение кислорода в артериальной крови. Напряжение углекислоты играет второстепенную роль, поскольку СО 2 легко отдается в водную среду. Вентиляция жабр служит для извлечения кислорода. Сигналы о содержании кислорода в воде поступают в дыхательный центр рыб из хеморецепторов жаберных сосудов - предшественников артериальных хеморецепторов воздушно-дышащих позвоночных. Переход животных к сухопутному образу жизни сопровождался с резким ограничением отдачи СО 2 через кожный покров - она стала выводиться почти исключительно легкими. И если напряжение кислорода в крови млекопитающих почти не изменилось по сравнению с рыбами, то напряжение СО 2 возросло почти в 12 раз, такое повышение обусловлено и затруднением выделения, и повышением интенсивности метаболических процессов в тканях. Все это потребовало развития мощных средств, обеспечивающих сохранение кислотно-основного гомеостаза внутренней среды организма. С другой стороны, требования к поддержанию гомеостаза в процессе эволюции стали более жесткими. Особое значение имеет прогрессивное развитие мозга, потому что нейроны отличаются высокой чувствительностью к химизму внеклеточной жидкости, в частности, к ее рН. В результате в ходе эволюции появились коренные изменения в регуляции дыхания. Гипоксический стимул уступил доминирующую роль гиперкапническому (это, безусловно, не изменило конечной цели функции дыхания: продукция СО 2 вполне может служить показателем кислородного запроса ткани). В этом легко убедиться, глядя на рисунок 14. На этом рисунке представлены изменения вентиляции легких (МОД в л/мин) в ответ на увеличение напряжения углекислого газа в крови – гиперкапнический стимул и в ответ на уменьшение напряжения кислорода в крови – гипоксический стимул. В связи с этим, основная функция в стимуляции дыхательного центра перешла от периферических хеморецепторов к центральным. Речь идет прежде всего о бульбарных хемочувствительных структурах, реагирующих на изменение концентрации ионов водорода и напряжения СО2 во внеклеточной жидкости мозга. За периферическими, артериальными хеморецепторами, которые возбуждаются и при повышении напряжения СО 2, и при снижении напряжения кислорода в омывающей их крови, осталась лишь вспомогательная роль в стимуляции дыхания. Поэтому рассмотрим сначала центральные хеморецепторы, которые оказывают более выраженное влияние на деятельность дыхательного центра.
Рисунок 14 Зависимость вентиляции легких
|
||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-26; просмотров: 670; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.222.119.143 (0.008 с.) |