Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Абсцисса — дни опытов (по М. Л. Пигаревой)Содержание книги
Поиск на нашем сайте
Хорошей экспериментальной моделью такой ситуации может служить выработка условнорефлекторного переключения разнородных рефлексов по Э. А. Асратяну, когда один и тот же сигнал (в опытах М. Л. Пигаревой — звук) утром подкрепляется пищей, а вечером — болевым раздражением. Двустороннее разрушение миндалин у крыс не позволяет добиться переключения на протяжении сорока дней (рис. 14). Выработка переключения оказалась возможной только при сочетании слабого болевого раздражения с высокой пищевой возбудимостью или, напротив, при сочетании кратковременной пищевой депривации с сильным болевым подкреплением оборонительного рефлекса. Многие дефекты условнорефлекторной деятельности и естественного поведения, возникающие у животных после повреждения миндалины, можно объяснить именно конкуренцией мотиваций. Крысы с повреждением корти-комедиальной области позже начинают есть в новой обстановке, поскольку их пищевая активность подавляется исследовательским поведением (Selafani, Belluzzi, Grossman, 1970). Нарушения пищевых и питьевых условных рефлексов у крыс после разрушения медиальной части миндалины становятся особенно сильными в случае кон-
Рис. 13. Процент оборонительных (А) и пищевых (Б) условных реакций у контрольных (/) и амиг-далоэктомированных (2) крыс (по М. Л. Пигаревой)
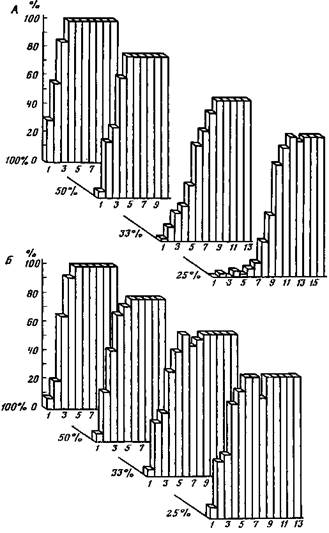
Рис. 14. Процент правильных оборонительных (/) и пищевых (2) условных реакций при выработке условнорефлектор-ного переключения у амигдалоэктомиро-ванной крысы (по М. Л. Пигаревой) куренции мотиваций, созданной определенной продолжительностью соответствующего депривирования (Korczyn-ski, Fonberg, 1976). Стимуляция миндалины у кошек ослабляет кожно-гальваническую реакцию на малозначимые звуковые сигналы (щелчки) и усиливает КГР на щелчки, сочетавшиеся с болевым раздражением (Гилин-ский, 1985). Повреждение вентромедиальной миндалины не нарушает пищевые инструментальные условные рефлексы и потребление пищи, но подавляет охотничье поведение кошки. Повреждение дорзолатеральной части дает противоположный эффект (Zagrodska, Fonberg, 1979). Разрушение дорзальной области не влияет на ловлю и поедание мышей при одиночном поведении кошки. Присутствие другого кота подавляет хищническую реакцию на появление мыши (Zagrodska, Brudnias-Ste-powska, Fonberg, 1983). Исследование пищедобыватель-ного поведения крыс в различных ситуациях привело авторов к выводу о том, что кортикобазомедиальная миндалина контролирует отношения между пищевой мотивацией и другими, сосуществующими с ней побуждениями (Lukaszewska, Korczynski, Kostarczyk, Fonberg, 1984). Таким образом, вывод о принадлежности миндалины к системе структур, определяющих выбор поведения, можно принять с уточнением, что миндалина участвует в этом выборе путем «взвешивания» конкурирующих эмоций, порожденных конкурирующими потребностями. Миндалина вовлекается в процесс организации поведения на сравнительно поздних этапах этого процесса, когда актуализированные потребности уже сопоставлены с перспективой их удовлетворения и трансформированы в соответствующие эмоциональные состояния. Прогнозирование вероятности удовлетворения потребности (вероятности подкрепления) осуществляется с участием преимущественно «информационных» мозговых структур — гиппокампа и передних отделов новой коры. Гиппокамп рассматривают как центр системы мозговых образований, моделирующих мир в трехмерном эвклидовом пространстве. При участии гиппокампа события, происходящие в окружающей среде, фиксируются во времени и пространстве: в гиппокампе свободно движущихся крыс найдены нейроны, кодирующие местоположение внешних объектов (Keefe, Nadel, 1979). Японским макакам красным светом показывали одну из двух кнопок. После периода задержки обезьяна должна была нажать на ранее показанную ей кнопку для того, чтобы получить порцию фруктового сока. Были выделены нейроны, изменявшие импульсную активность 1) во время освещения кнопки, 2) в период задержки, 3) в момент выбора кнопки, 4) при нажатии на кнопку и 5) при подкреплении или его отсутствии (Watanabe, Niki, 1985). Полагают, что основной функцией гиппокампа является «рабочая» (свежая, промежуточная и т. п.) память независимо от того, идет ли речь о фиксации пространственного или временного материала (Olton, Becker, Handelmann, 1979), причем эти функции сходны у человека и высших животных (Sahgal, 1980). Гиппокамп вовлекается в процесс обучения на самых ранних его этапах (Segal, Olds, 1972). При выработке пищевых условных рефлексов на звук коротколатентные ответы нейронов были зарегистрированы в гиппокампе, а длиннолатентные условные ответы — в височной коре. По мнению авторов, гиппокамп является первым пунктом конвергенции условных и безусловных возбуждений. Именно в гиппокампе и латеральном отделе перегородки были найдены клетки, активность которых изменялась толькго при спаренных стимулах. Заметим, что разрушение дорзального и вентрального гиппокампа у крыс не нарушает их способность к обучению и даже облегчает выработку условнорефлекторного переключения (Пигарева, 1973). На рис. 15 можно видеть, что после десяти дней безуспешных попыток выработать переключение оборонительного и пищевого условных рефлексов у трех крыс двусторонняя гиппокампэктомия (рис. 16) на протяжении двух недель привела к формированию стабильного условнорефлекторного переключения. Наиболее ярким дефектом гиппокампэктомированных животных оказалась их чувствительность к ситуациям с низкой вероятностью подкрепления условных сигналов (Kimble, Kimble, 1970; Jarrard, Becker, 1977; Devenport, Hollowy, 1980). По данным М. Л. Пигаревой (1978), при вероятности подкрепления пищевых условных рефлексов, равной 100 и 50%, гиппокампэктомированные крысы хотя и отстают от интактных, но все же справляются с задачей. Выработка условных рефлексов при вероятности 33 и 25% оказалась для них недоступной (рис. 17). Выпадение реакций на сигналы с низкой вероятностью подкрепления ведет к тому, что гиппокампэктомированные крысы превосходят контрольных в различении сигналов с разной вероятностью их подкрепления (Means, Walker, Isaacson, 1970; Stevens, 1973), частным случаем чего
Рис. 15. Динамика выработки условнорефлекторного переключения у трех крыс до и после двустороннего повреждения гиппокампа (показано стрелкой^ / — оборонительные реакции; 2 — пищевые (по М. Л. Пигаревой)
Рис. 16. Реконструктивная схема двустороннего разрушения гиппокампа Заштрихован максимальный объем повреждения, черным цветом показан минимальный объем
Рис. 17. Процент правильных условных пищевых реакций у контрольных (/) и гиппокампэктомированных (2) крыс при различной вероятности подкрепления условного сигнала Абсцисса — дни опытов (по М. Л. Пигаревой) является облегчение у этих животных выработки услов-норефлекторного переключения. Нарушения эмоциональной сферы при повреждении гиппокампа, как правило, незначительны (Карамян, Сол-лертинская, 1982). Вместе с тем стимуляция дорзально-го гиппокампа тормозит эмоциональные реакции собак, вызванные прямым раздражением гиппокампа (Петров, 1974), а стимуляция вентрального гиппокампа кролика подавляет самораздражение латерального гипоталамуса (Koreli, 1977). Повреждение вентрального гиппокампа кролика извращает эффекты прямой электрической стимуляции медиального гипоталамуса: эмоционально отрицательные реакции избегания сменяются самораздражением тех же самых структур (Давитулиани, Корели, 1982). Таким образом, гиппокамп оказывает регулирующее влияние на эмоциогенные структуры гипоталамуса. О функциональных связях гиппокампа с гипоталамусом свидетельствует и тот факт, что повреждение заднего гипоталамуса изменяет электрическую активность гиппокампа в стадии быстрого сна (Гветадзе, 1980). Будучи структурой, где мотивационное возбуждение от заднего и переднелатерального гипоталамуса сопоставляется с информацией, поступающей из внешней среды (через перегородку), равно как и со следами ранее накопленного опыта (из коры), гиппокамп, по-видимому, осуществляет двойную функцию. Во-первых, он играет роль входного фильтра информации, подлежащей или не подлежащей регистрации в долговременной памяти (Виноградова, 1975). С другой стороны, гиппокамп участвует в извлечении следов из памяти под влиянием мотивационного возбуждения для использования этих следов в организации текущего поведения (Hirsh, 1974). Метод корреляционного анализа электрической активности позволил установить, что на ранних стадиях выработки условного рефлекса тета-волны в гиппокампе опережают тета-волны височной коры, т. е. импульсы возбуждения следуют из гиппокампа в кору. После упрочения условной связи тета-волны энторинальной коры опережают волны в гиппокампе, импульсы возбуждения распространяются из коры в гиппокаип (Adey, Walter, Hendrix, 1961). Подчеркнем, что следы могут быть извлечены из памяти независимо от внешних стимулов и обеспечить ожидание этих стимулов по механизму тонических обратных условных связей (Асратян, 1974). Электрофизиологическим коррелятом механизма, квантующего поток извлекаемых из памяти энграмм, является тета-ритм, столь характерный для электрической активности гиппокампа. Все ситуации, в которых мы наблюдаем усиление тета-ритма, будь то ориентировочный рефлекс, поисковое поведение, организация сложных неавтоматизированных движений, появление признаков эмоционального напряжения и т. п., обладают одной общей для них чертой: перечисленные случаи требуют активной мобилизации ранее выработанных условных связей, извлечения хранящихся в памяти энграмм для сопоставления с поступающими извне сигналами или для пересмотра, рекомбинации следов в целях построения новых приспособительных действий (Gray, 1982). Изложенное выше позволяет говорить о важной роли гиппокампа в творческой деятельности мозга, в порождении гипотез, тем более что у человека гиппокамп доминантного полушария вовлекается в анализ словесных сигналов, а гиппокамп правого полушария — в анализ невербальных стимулов. Подобная функциональная асимметрия противоречит представлению о гиппокампе как о древнем и потому примитивном образовании, способном только к осуществлению элементарных функций. По мнению А. С. Батуева (1981), мотивационное возбуждение распространяется из лимбической системы в лобную и частично в теменную кору. В сущности, лобную область можно рассматривать как неокортикальное продолжение лимбической системы (Nauta, 1964). Сведения о внешней среде поступают в ассоциативные системы неокортекса через ассоциативные ядра таламуса и транскортикальным путем от сенсорных проекционных зон. На основе этой информации и извлеченных из памяти энграмм фронтальная кора строит вероятностный прогноз, а таламопариетальная система обеспечивает селективное внимание к значимым факторам внешней среды и к ведущим звеньям предстоящего двигательного акта. Иными словами, теменная кора осуществляет приурочи-вание двигательных реакций к пространственной организации внешнего мира. Решающее значение передних отделов новой коры для прогнозирования вероятности предстоящих событий установлено у крыс, кошек, собак, обезьян и человека. Нарушение вероятностного прогноза после повреждения лобной коры сохраняется у обезьян в течение трех с половиной лет. Двустороннее повреждение теменной коры вызывает нарушение этой функции с быстрым (на протяжении одного — полутора месяцев) ее восстановлением (Маликова, 1981). Повреждение префронтальной ассоциативной коры у кошек нарушает предпочтение кормушки с высокой вероятностью подкрепления, в то время как удаление теменной коры не дает такого дефекта (Батуев, Куликов, 1983). Возможно, что именно нарушением механизмов вероятностного прогнозирования объясняется отвлекаемость больных на малозначимые для них стимулы при опухолях лобных долей мозга (Белый, 1979). Вместе с тем вероятностная структура среды получает отражение в активности нейронов и ряда других областей неокортекса. В опытах Д. Г. Шевченко (1982) кролик в ответ на вспышку справа или слева нажимал на педаль и получал пищу из соответствующей кормушки. Вспышки подавались с двух сторон с вероятностью 100 и 50%. Реакция нейронов зрительной коры отличалась в зависимости от вероятности появления вспышки перед нажатием на педаль и не отличалась на других этапах поведения. Подчеркнем, что в отличие от гиппокампа, сохранность которого необходима для поддержания реакций на сигналы с низкой вероятностью подкрепления, фронтальная кора важна для ориентации поведения на сигналы высоковероятных событий. Так, после удаления фронтальных отделов неокортекса у крыс и собак значение часто и редко подкрепляемых условных сигналов как бы уравнивается: все сигналы становятся в одинаковой мере эффективными (Андреев, 1969; Пигарева, 1983). На рис. 18 показана динамика выработки пищевых условных рефлексов у крыс при различном подкреплении до (рис. 18, Л) и после разрушения передних отделов новой коры (рис. 18, Б; 19). Видно, что процесс формирования условного рефлекса замедляется при высокой вероятности подкрепления и ускоряется — при низкой. Иными словами, сигналы с различной вероятностью их подкрепления пищей становятся одинаково эффективными. Этот результат представляет тем больший интерес, что, судя по имеющимся данным, фронтальные области коры головного мозга крыс не отличаются по своим основным функциям от фронтальной коры высших позвоночных, включая приматов (Kolb, 1984). В процессе экспериментов с вероятностным прогнозированием В. М. Русалов обнаружил три группы обследованных им лиц. Представители первой группы адекватно
Рис. 18. Процент двигательных пищевых условных реакций у контрольных (А) и лобэктомированных (Б) крыс при различной вероятности подкрепления Абсцисса — дни опытов (по М. Л. Пигаревой) 5 П. В. Симонов
Рис. 19. Реконструктивная схема удаления фронтальных областей неокортекса у крыс отражают характеристики вероятностной среды (вероятностное соответствие). Представители второй группы занижают частоту более частого события (вероятностное безразличие), а представители третьей — завышают ее (стратегия максимализации). Эксперименты на животных с последовательным выключением мозговых структур позволяют предположить, что субъекты первой группы обладают хорошо сбалансированными функциями лобной коры и гиппокампа, у лиц второй группы относительно преобладают функции лобвой коры, а у представителей третьей — функции гиппокампа. В результате субъективная оценка вероятности событий у представителей последних двух групп не соответствует объективной вероятности редких и частых событий (Русалов, 1979). Эффективность совместного прогнозирования лицами, принадлежащими к противоположным группам, скорее подкрепляет высказанное предположение, чем противоречит ему (Русалов, 1982). В литературе имеются сведения о том, что вероятностная структура среды более адекватно отражается правым полушарием мозга человека, в то время как левое полушарие одинаково реагирует на сигналы с высокой и низкой вероятностью (Крауклис, Янсон, Ширяев, Козановская, 1979). Способность фронтальной коры производить селекцию высокозначимых сигналов, отсеивая второстепенные для доминирующей мотивации стимулы, возможно, осуществляется с участием хвостатого ядра, чьи нейроны вовлекаются в процесс выработки тормозных условных рефлексов быстрее, чем в реакции на положительные сигналы (Суворов, Данилова, Дрягин, Шуваев, 1981). Отметим, что у собак нарушение вероятностного прогнозирования наблюдается после удаления дорзолатераль-ных, а не медиобазальных областей префронтальной коры (Мехедова, 1974). Таким образом, в пределах лобного неокортекса обнаруживается преимущественная причастность дорзолатеральных отделов к «информационной» системе мозговых образований, а медиобазальных отделов—к «мотивационной» системе. Такому разделению функций хорошо соответствуют анатомические связи этих двух частей лобной коры: дорзальной части — с гип-покампом, а вентральной — с миндалиной (Nauta, 1972). Лобная кора может влиять на гиппокамп через пучок волокон к поясной извилине. Префронтальная кора — единственная неокортикальная область, которая получает иннервацию из вентральной части покрышки и миндалины (Divac, Kosmal, 1978). Исследования показали, что нейронная активность дорзолатеральнои префронтальной коры обезьян зависит от интеграции сенсорных процессов, необходимых для организации поведения, в то время как нейроны орбитофронтальной коры связаны с мотивацией, их активность зависит от качества подкрепления (Aou Shuji, 1982). Реакции некоторых из этих нейронов чрезвычайно специализированны: они отвечают на апельсин, на змею, на паука и т. д., хотя большинство нейронов избирательно реагирует только в зависимости от пищевых или аверсивных качеств стимула (Thorpe, Rolls, Maddison, 1983). Мотивирующие влияния поступают во фронтальную кору из латерального гипоталамуса (Oomura, Fakiga-ma, 1976). По данным Э. Роллса и С. Купера (Rolls, Cooper, 1973), в отличие от других областей неокортекса нейроны префронтальной коры крыс реагировали на стимуляцию тех структур мозга, которые вызывают феномен самостимуляции. В префронтальной коре имеются нейроны, которые у голодных обезьян отвечают на появление пищи в кормушке до начала пищедобывательного движения (Орлов, Пирогов, Шефер, 1979). Активность этих нейронов ослабевает по мере насыщения, но 5* вновь возникает при замене пищи на более вкусную (Батуев, 1985). При исследовании инструментальных пищевых отсроченных условных реакций у обезьян обнаружены: 1) нейроны моторных программ, реагирующие на открытие экрана, которое служит сигналом начала движения; 2) нейроны сенсорных программ, разряжающиеся во время отсрочки; 3) нейроны подкрепления, чья активность изменяется при появлении пищи (Батуев, 1984). Во фронтальной коре зарегистрированы нейроны, которые реагируют на отсутствие подкрепления при осуществлении инструментальной реакции, причем эти нейроны не отвечают на отсутствие подкрепления в ситуации классического обусловливания, когда появление пищи не зависит от действий обезьяны. Нейроны ошибочных реакций локализованы в медиовентральных зонах префронтальной коры (Niki, Hiroaki, 1982). Таким образом, передние отделы новой коры оказываются причаст-ны и к программированию поведенческого акта, и к оценке его результатов. Нейроны ростральной дорзолатераль-ной префронтальной коры преимущественно участвуют в процессах мотивации, а нейроны каудальной части — в сенсорном анализе вкуса и качества пищевого подкрепления (Inoue, Oomura, Aou, Nishino, Sikdar, 1985). Первые сдвиги клеточной активности при выработке нового условного рефлекса отмечены во фронтальной, сенсомоторной коре и в гиппокампе. Нейроны сенсомо-торной коры реагируют через 20 мс после звукового условного сигнала, подкрепляемого миганием, т. е. одновременно с нейронами первичной слуховой коры (Батуев, 1984). Наличие первичных пластических перестроек в нейронах новой коры побуждает некоторых авторов различать два механизма выработки навыков, один из которых связан, а второй не связан с лимбической системой. Последний механизм формируется у обезьян к 3—4 месяцам, в то время как система памяти с участием лимби-ческих структур созревает медленно и не достигает уровня взрослых даже через год после рождения (Bacheva-lier, Mishkin, 1984). Следует различать «лимбическую» память как активное извлечение информации, необходимой для удовлетворения потребности, и феномен напоминания, ограничивающийся выбором знакомого стимула из ряда предъявленных. Последний механизм не страдает до тех пор, пока сохранена височная кора (Malamut, Saunders, Mishkin, 1984). Анализируя функции неокортекса, необходимо учитывать их латерализацию, наметившуюся уже у животных и испытывающую влияние раннего индивидуального опыта. У цыплят, певчих птиц, крыс и приматов левое полушарие оказалось сильнее вовлечено в процессы коммуникации, а правое ответственно за ориентировку в пространстве и эмоции (Denenberg, 1981). Исследование восприятия значимых внутривидовых сигналов у японских макак обнаружило преимущество реакций на сигналы, подаваемые в правое ухо. Подача сигналов, принадлежащих другому виду, латерализации не выявила (Peterson, 1978, 1984). Отсасывание коры правой лобной доли у крыс вызывает их гиперактивность и падение уровня норадреналина в коре обоих полушарий. Экстирпация левой лобной доли таких изменений не дает (Pearl-son, Robinson, 1981). По данным В. Л. Бианки (1985), к функциям правого полушария у крыс следует отнести восприятие пространства и конкретных признаков внешних объектов, одновременную обработку информации, явления дедукции. К функциям левого полушария — восприятие времени и абстрактных признаков, последовательную обработку информации, явления индукции. Стадия генерализации условных рефлексов теснее связана с деятельностью правого полушария. Динамический стереотип лучше воспроизводится левым. Интересно, что у человека левое полушарие активировано сильнее при первых реализациях задания, связанного, например, с мысленным воспроизведением значимых ситуаций. По мере повторения этой процедуры возникает асимметрия уровня активации в пользу правого полушария (Русало-ва, 1983). Завершая обзор функций передних отделов новой коры, гиппокампа, миндалины и гипоталамуса, мы приходим к выводу о том, что взаимодействие этих четырех структур оказывается необходимым и достаточным для организации поведения в системе координат «потребности— вероятность их удовлетворения в окружающей внешней среде». Все остальные образования мозга играют исполнительную или вспомогательную роль, будь то сенсорные системы, механизмы построения движений (пирамидные и экстрапирамидные), системы регуляции уровня бодрствования и вегетативных функций. Что касается других структур лимбической системы, то перегородка настолько тесно объединена с гиппокампом, что большинство исследователей предпочитают говорить об единой септо-гиппокампальной системе (Gray, 1982). Центральное серое вещество специфически связано с оценкой аверсивности стимулов, с организацией реакций избегания, например, при раздражении медиального гипоталамуса (Sandner, Schmitt, Karli, 1982). Неспецифический таламус служит местом конвергенции сенсорных и мотивационных импульсов (Casey, Keene, 1973), и его значение не выходит за пределы передаточно-релейных функций. Хотя в таламусе обезьян обнаружены нейроны, частота разрядов которых уменьшается или возрастает и стойко удерживается при выработке условных рефлексов, большой скрытый период (200 мс после предъявления условного сигнала) позволяет рассматривать эти реакции как вторичный отзвук событий, разыгрывающихся в каких-то других образованиях мозга. Иными словами, именно перечисленные выше четыре структуры определяют, на какие внешние стимулы и какой поведенческой реакцией в каждый данный момент ответит живой организм. По мнению Могенсона с соавторами, связь лимбической системы с двигательной осуществляется через вентральный отдел покрышки среднего мозга и прилегающее ядро с бледным шаром (Mogenson, Jones, Yim Chi Yiu, 1980). Разрушение прилегающего ядра увеличивает латентный период реакции избегания у крыс и снижает их стремление к доминированию, не влияя на двигательную активность в открытом поле и пищедобы-вательное поведение (Fantin, Bottecchia, 1984). Что же касается согласованной деятельности «принимающих решение» структур, то к ним в полной мере относится глубокая мысль А. А. Ухтомского о том, что «соподчинение физиологических приборов в порядке их нервной увязки есть процесс вынужденный и не предполагает для себя вмешательства какого-либо дополнительного, специального „координирующего центра"» (Ухтомский, 1954, с. 127). Последовательно рассмотрев функциональную специализацию отдельных мозговых структур, попытаемся представить динамику их взаимодействия в целом (рис. 20). Внутренние (метаболизм) и внешние (боль, запах и т. д.) побуждающие безусловные стимулы активируют мотивационные структуры гипоталамуса, который, в свою очередь, активирует гиппокамп и передние отделы новой коры. Благодаря гиппокампу широкий круг внешних стимулов усиливает доминантное состояние.
Рис. 20. Схема взаимодействия мозговых структур в процессе организации поведенческого акта Внутренние (метаболизм) и внешние (боль, запах и т. д.) побуждающие безусловные стимулы активируют мотивационные структуры гипоталамуса (/Т), который, в свою очередь, активирует гиппокамп {ГИП) и передние отделы новой коры (влияние гипоталамуса на миндалину выражено слабее). Благодаря гиппокампу широкий круг внешних стимулов усиливает доминантное состояние. В случае совпадения этих стимулов с действием подкрепляющих безусловных раздражителей гиппокамп оказывается первым местом встречи сочетаемых афферентаций. При сформированном поведенческом акте в результате совместной деятельности гиппокампа и фронтальной коры (ФК) отбираются те внешние стимулы или их энграммы, которые ранее сопровождались удовлетворением данной потребности. Путем сопоставления мотивационного возбуждения с наличными стимулами и с энграммами, извлеченными из памяти, в миндалине (М) формируется эмоциональная окраска этих стимулов и энграмм, что ведет к выделению доминирующей мотивации, подлежащей первоочередному удовлетворению. Сложившаяся во фронтальной коре программа действия поступает в базальные ганглии, где путем взаимодействия с теменной корой «вписывается» в пространственные координаты предстоящего действия. Отсюда через моторную кору возбуждение поступает на эффекторные органы, реализующие целенаправленное поведение. Сплошные линии — информационная афферентация, прерывистые — мотивационные влияния, двойные — эмоционально окрашенная афферентация В случае совпадения этих стимулов с действием подкрепляющих безусловных раздражителей гиппокамп оказывается первым местом встречи сочетаемых афферентаций. При сформированном поведенческом акте в результате совместной деятельности гиппокампа и фронтальной коры отбираются те внешние стимулы или их энграммы, которые ранее сопровождались удовлетворением данной потребности. Путем сопоставления мотивационного возбуждения с наличными стимулами и энграммами, извлеченными из памяти (через височную кору?), в миндалине формируется эмоциональная окраска этих стимулов и энграмм, что ведет к выделению доминирующей мотивации, подлежащей первоочередному удовлетворению. Сложившаяся во фронтальной коре программа действия поступает в стриатум, где путем взаимодействия с теменной корой «вписывается» в пространственные координаты 'Предстоящего действия. Регистрация вызванных потенциалов показала, что на стадии формирования условного рефлекса теменная кора связана преимущественно с кортикальным представительством сенсорных систем, а по мере стабилизации — с моторной корой (Попова, Гнеушев, Деревягин, 1982). Изменения нейронной активности полосатого тела, бледного шара и черной субстанции предшествуют движениям и возникают в ответ на внешние сенсорные стимулы только в тех случаях, где эти стимулы и движения имеют поведенческое значение (Evarts, Kimura, Wurtz, Hikosaka, 1984). Авторы заключают, что инициация двигательных актов опосредована активностью базальных ганглиев и связанной с ними фронтальной корой. Импульсация из базальных ганглиев и мозжечка приходит в кору через таламус. В ответ на сигнал к движению зубчатые нейроны мозжечка разряжаются через 30 мс, а нейроны коры через 40 мс после сигнала. Нигростриальные пути участвуют в подготовке и регуляции движений, но не в их мотивации (Rolls, Rolls, 1982). Из фронтостриатной системы возбуждение через моторную кору поступает на эффек-торные органы, реализующие целенаправленное поведение. Разумеется, предлагаемая схема организации поведения носит крайне обобщенный характер, нуждается в дальнейшем уточнении и экспериментальном подтверждении применительно к различным видам высших млекопитающих. Вместе с тем она дает возможность рассмот- реть интегративную деятельность мозга в целом, не противореча имеющимся фактическим данным и анатомически прослеженным связям между структурами головного мозга. Мы можем обсуждать эту деятельность, оставаясь на физиологической почве и не прибегая к понятиям, заимствованным из других областей знания, типа «моделирование внешнего мира», «принятие решения», «рассудочная деятельность» и т. д. и т. п. С другой стороны, уточнив роль различных мозговых структур в организации поведения, мы приобретаем возможность рассматривать индивидуальные особенности поведения человека и высших животных как результат индивидуальных — врожденных и приобретенных — особенностей взаимодействия этих структур. Подобный подход к проблеме индивидуальных (типологических) различий был в свое время намечен И. П. Павловым и заслуживает дальнейшей экспериментальной и теоретической разработки.
|
||||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 220; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.21.105.46 (0.014 с.) |