Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Диссоциация рибосомы – необходимая предпосылка для инициации.Содержание книги
Поиск на нашем сайте По окончании синтеза молекулы белка рибосомные субъединицы диссоциируют на составляющие малую и большую субъединиы. Это состояние подерживается специальными белками, которые связываются со свободной малой рибосомной субъединицей [30S у прокариот или 40S у эукариот] и препятствуют ее взаимодействию с большой субъединицей. Эти белки называют факторами инициации или IF [Initiation Factors].Таких белков у прокариот три, а у эукариот их значительно больше. Их принято обозначать как IF1, IF2, IF3 у прокариот, в названии факторов эукариот прибавляется буква е (eIF-1, eIF-2, eIF-3, eIF-4A, eIF-4B, eIF-4C, eIF-4D, eIF-4E, eIF-4F, eIF-5 и т.д.). Функции отдельных факторов приведены в таблицах
Наибольшим сродством к малой субъединице рибосомы обладает IF3, котрорый является ведущим белком, поддерживающим эту субъединицу в свободном нативном состоянии. Другой фактор, IF2, является представителем большой группы белков, способных связываться с гуаниловыми нуклеотидами и обладающими ГТФазной активностью. Он вступает в процесс на следующем этапе инициации, который может осуществляться двумя способами а) нативная малая субъединица вначале взаимодействует с иРНК, и затем к этому комплексу присоединяется инициирующая аминоацил-тРНК связанная с IF2 и ГТФ б) с малой субъединицей вначале взаимодействует инициирующая аминоацил-тРНК с IF2 и ГТФ, а затем к этому комплексу присоединяется иРНК
Рис..Образование инициирующего комплекса у прокариот.
Как видно и тот и другой способы приводят к образованию так называемого инициирующего комплекса, состоящего из малой рибосомной субъединицы, иРНК, инициирующей аминоацил-тРНК и факторов инициации. Образование инициирующего комплекса решает важную задачу точной установки инициирующей аминоацил тРНК в начале кодирующей последовательности иРНК.
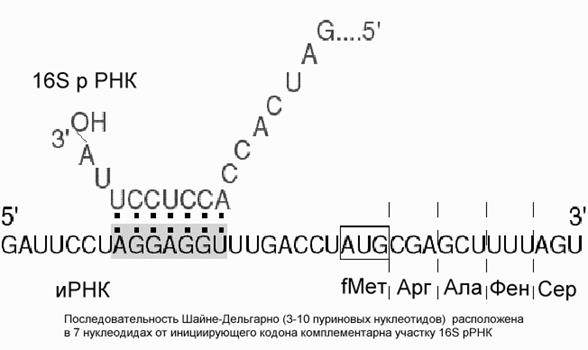
Рис. Механизм взаимодействия иРНК и рибосомы у прокариот. Основная роль в этом взаимодействии принадлежит комплементарному связыванию 16S РНК рибосомы и специфической последовательности на иРНК (последовательность Шайне-Дельгарно)
У прокариот и у эукариот это происходит по-разному. У прокариот малая рибосомная субъединица с факторами инициации имеет повышенное сродство к специальному внутреннему участку иРНК, который называют инициирующим, или рибосомосвязывающим участком.. Наиболее известной универсальной структурой такого участка является полипуриновая нуклеотидная последовательность Шайне-Дельгарно, отстоящая от инициирующего кодона на несколько (3-10) нуклеотидов к 5'-концу, т.е. предшествующей инициирующему кодону. Эта последовательность в большей или меньшей степени комплементарна 3'-концевому участку рибосомной 16S РНК, богатому пиримидиновыми нуклеотидными остатками (5'-GAUCACCUCCUUA-3' у E.coli). Комплементарными могут быть от 3 до 9 остатков (обычно около 4-5). Последовательность CCUCC 3'-концевого участка 16S РНК E.coli оказывается наиболее часто комплементарной (хотя, как правило, частично) полипуриновой прединициирующей последовательности иРНК. Считается, что комплементарное спаривание 3'-концевого участка 16S РНК с прединициирующим полипуриновым блоком иРНК и обеспечивает точную установку инициирующего комплекса на начало кодирующей последовательности иРНК. У эукариот малая рибосомная субъединица с факторами инициации узнает кэп 5'-концевого участка иРНК, и как правило, взаимодействует с ним. У эукариот малая субъединица обычно взаимодействует с инициирующей аминоацил-тРНК до связывания иРНК. Связавшись с 5'-концом иРНК вместе с инициирующей аминоацил-тРНК и факторами инициации, малая рибосомная субъединица начинает двигаться от 5'-конца по направлению к 3'-концу, используя энергию гидролиза АТФ для раскручивания вторичной структуры иРНК. АТФазной и хеликазной активностью обладает специальный эукариотический фактор инициации eIF4. Когда рибосомная частица достигает и узнает инициирующий кодон, антикодон инициирующей аминоацил-тРНК взаимодействует с ним, обеспечивая тем самым узнавание начала кодирующей последовательности иРНК.
И у прокариот и у эукариот инициирующей аминоацил-тРНК является метионил-тРНК, у прокариот, как уже отмечалось, ее аминогруппа блокирована формильным остатком -формил-метионил-тРНК. Ее антикодон всегда ЦАУ, так что он полностью комплементарен кодону АУГ и частично комплементарен другим возможным инициаторным кодонам прокариот [например, ГУГ и УУГ]. Структура молекулы тРНК выпонящей роль инициирующей метионил-тРНК несколько отличается от структур других тРНК, в том числе и от структуры метионил-тРНК, используемой в процессе элнгации, что и делает ее функцию в качестве инициатора трансляции уникальной. Инициаторные тРНК имеют консервативный участок ГАУЦ в положении 51-54. Универсальная последовательность в тех же позициях элонгаторных тРНК содержит T вместо A и псевдоуридин вместо У. В инициаторных тРНК дигидроуридиловая петля не содержит дигидроуридина Особенности структуры инициирующей метионил-тРНК обеспечивают ее специфическое комплексирование с белком IF2 при наличии ГТФ [ГТФ, присоединяясь к белку, повышает сродство белка к инициирующей метионил-тРНК]. Универсальность инициации с участием инициирующей метионил-тРНК приводит к тому, что всегда и
Рис..Схема механизма инициации трансляции у эукариот.
везде любая полипептидная цепь, синтезируемая на рибосоме, начинается с метионина. Лишь потом, в ходе трансляции или после нее, этот концевой метионин может отщепиться специальной протеазой, что и происходит в большинстве случаев
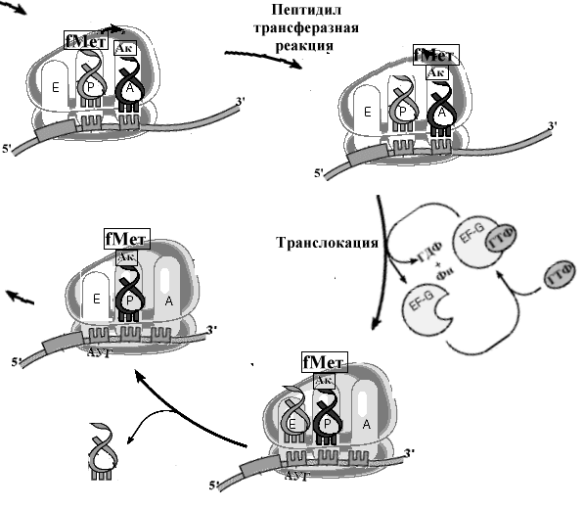
Рис. Схема реакций механизма элонгации процесса трансляции.
На этом этапе в процесс включается свободная большая рибосомная субъединица. Взаимодействуя с малой субчастицей и фактором IF2, большая суъединица активирует ГТФазную активность IF2, в результате чего ГТФ гидролизуется до ГДФ и ортофосфата, что приводит к снижению сродства IF2 к инициирующей метионил-тРНК и IF2 с ГДФ легко вытесняются из рибосомы. На большой субъединице формируется пептидильный участок (Р участок), на котором размещается инициирующая метионил-тРНК. Вытесняются одновременно и все другие факторы инициации, включая IF3. В итоге образуется полная 70S [ у прокариот] или 80S [ у эукариот] рибосома с Р-участком, занятым инициирующей метионил-тРНК, и с вакантными А- и Е участками. Этот завершающий шаг процесса инициации начинает образование и элонгацию пептида. На вакантный А-участок поступает первая аминоацил-тРНК в комплексе с фактором элонгации EF1(EF-Tu у прокариот) и ГТФ. После гидролиза ГТФ, EF1(EF-Tu) с ГДФ покидают рибосому, а аминоацил-тРНК, связанная на А-участке, получает возможность реагировать с инициирующей метионил-тРНК в реакции транспептидации, катализируемой большой субъединицей рибосомы. Фактор EF1 (EF-Tu) вновь подключается в процесс трансляции после замены ГДФ на ГТФ при участии факторов, стимулирующих этот обмен гуаниловых нуклеотидов. Так начинается элонгация.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 455; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.62 (0.009 с.) |


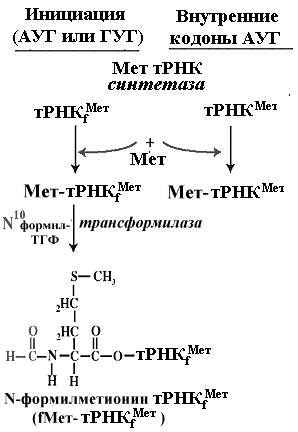 Единственным инициирующим кодоном эукариот является триплет АУГ. У прокариот с этого триплета также начинается 90% всех кодирующих последовательностей. Однако некоторая часть прокариотических мРНК инициирует трансляцию на других триплетах - ГУГ, реже УУГ и совсем редко на некоторых других. Очевидно, что у прокариот способность некоторых триплетов быть инициирующими определяется их положением в инициирующем или рибосомосвязывающем участке иРНК. Эти же триплеты, когда они встречаются в кодирующей последовательности в ходе элонгации, кодируют ту или иную аминокислоту АУГ - метионин, ГУГ - валин, УУГ - лейцин.
Единственным инициирующим кодоном эукариот является триплет АУГ. У прокариот с этого триплета также начинается 90% всех кодирующих последовательностей. Однако некоторая часть прокариотических мРНК инициирует трансляцию на других триплетах - ГУГ, реже УУГ и совсем редко на некоторых других. Очевидно, что у прокариот способность некоторых триплетов быть инициирующими определяется их положением в инициирующем или рибосомосвязывающем участке иРНК. Эти же триплеты, когда они встречаются в кодирующей последовательности в ходе элонгации, кодируют ту или иную аминокислоту АУГ - метионин, ГУГ - валин, УУГ - лейцин.