Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Тип бластулы зависит от типа дробления.Содержание книги
Поиск на нашем сайте
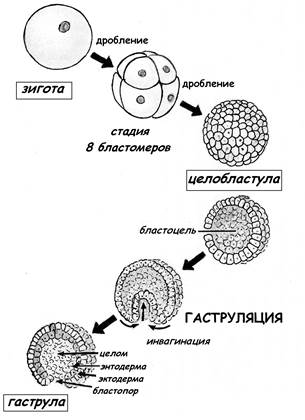
При полном равномерном синхронном дроблении первично изолецитальной яйцеклетки Ланцетника образуется целобластула ( рис.8): • стенка целобластулы – бластодерма образована одним слоем клеток (однослойная); • бластомеры примерно одинакового размера; • в середине целобластулы есть полость – бластоцель.
Рис.8. Ранние стадии развития ланцетника
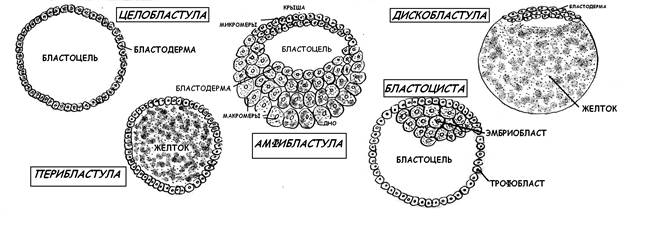
В результате полного неравномерного дробления мезолецитальной яйцеклетки амфибий (лягушки) образуется амфибластула (рис.9) : • на анимальном полюсе, которое содержало небольшое количество желтка, клеточные деления происходят с большой скоростью и образуются мелкие бластомеры – микромеры; • на вегетативном полюсе, богатом желтком, клетки делятся медленно и образуются крупные бластомеры – макромеры; • бластодерма (стенка бластулы) многослойная; • бластоцель смещён к анимальному полюсу; • анимальный полюс с многочисленными микромерами называется крышей бластулы, вегетативный с с макромерами образует дно; между ними краевая зона.
Рис.9. Ранние стадии эмбриогенеза амфибий Тип дробления у птиц с полилецитальной резко телолецитальной яйцеклеткой - неполное (частичное) дискоидальное: деление ограничено небольшим участком на анимальном полюсе, остальная часть желтка остаётся нераздробленной. В результате такого типа дробления образуется дискобластула (рис.10): • дискобластула имеет форму круглой пластинки (диска) на анимальном полюсе; бластодерма многослойная; • щелевидный бластоцель находится между бластодермой и синцитиальном слоем на поверхности желтка.
Рис.10. Типы бластул Характер дробления вторично изолецитальной яйцеклетки человека – полное неравномерное асинхронное. Образующая бластула на ранней стадии носит название морула (до стадии 64 бластомеров), а после появления полости – бластоцеля – бластоциста (рис.10). ГАСТРУЛЯЦИЯ Гаструляция – это следующий за дроблением период развития зародыша. Скорость клеточных делений снижается, интерфаза между митозами удлиняется, а сами клетки начинают активно синтезировать РНК и белки. Главная особенность этого периода – направленные активные перемещения делящихся клеток (клеточных потоков), которые приводят к глубокой перестройке зародыша и в конечном итоге к образованию трёх зародышевых листков: • эктодерма – наружный зародышевый листок; • энтодерма – внутренний зародышевый листок; • мезодерма - средний зародышевый листок, лежащий между экто- и энтодермой Способы гаструляции у животных разных систематических групп сильно варьируют. Описано четыре основных типа морфогенетических движений гаструляции (рис.11): • инвагинация; • эпиболия; • деламинация; • иммиграция
Рис.11. Основные механизмы гаструляции
Инвагинация – это впячивание бластодермы вегетативного полюса бластулы в полость бластоцеля, при этом образуется первичная кишка с полостью – гастроцелем, который сообщается с наружной средой отверстием – бластопором (первичным ртом). Инвагинация происходит при гаструляции у примитивных хордовых, таких как Ланцетник (рис.8). Эпиболия – перемещение активно делящихся мелких клеток (микромеров) анимальной части бластодермы в направлении вегетативного полюса. Микромеры обрастают крупные клетки (макромеры) вегетативного полюса, и макромеры погружаются внутрь зародыша. Эпиболия, как тип движений гаструляции, характерна для эмбриогенеза амфибий. Деламинация – расщепление (расслоение) бластодермы на два пласта: наружный – эпибласт и внутренний – гипобласт. Иммиграция – перемещение отдельных клеток из бластодермы внутрь зародыша. Если небольшие участки определенной области поверхности бластулы окрасить прижизненным красителем, можно проследить дальнейшую судьбу, дифференцировку клеток этого участка. Экспериментально показано, что клетки анимального полюса – клетки будущей эктодермы. По результатам таких исследований можно составить карту презумптивных зон. ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ ПТИЦ Тип яйцеклетки у птиц – полилецитальная, резко телолецитальная яйцеклетка. Тип дробления зиготы птиц- неполное (частичное, меробластическое) дискоидальное. Дробление ограничено лишь малой частью зиготы на анимальном полюсе (дробится лишь 1/500 часть зиготы), остальная часть зиготы не подвергается делению и представляет собой нераздробившийся желток. При дроблении возникают бластомеры различной величины, и в конечном итоге формируется дискобластула: бластомеры располагаются в форме диска, лежащего на массе нераздробившегося желтка (рис.10). Центральная часть дискобластулы называется зародышевым диском (бластодиском). Гаструляция в дискобластуле происходит в две фазы:
Первая фаза гаструляции- деламинация - заключается в расщеплении клеточной массы бластодиска на 2 листка: гипобласт (первичная энтодерма) и эпибласт (первичная эктодерма ) (Рис.12).
Рис.12. Деламинация в эмбриогенезе птиц
4. нераздробившийся желток.
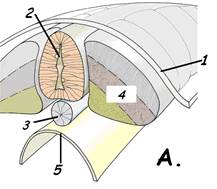
Клетки внутреннего листка - гипобласта - смыкаются в правильную однослойную пластинку. Наружный слой бластодиска – эпибласт – представляет собой клеточный пласт из более высоких призматических клеток. Центральный утолщенный участок бластодиска, состоящий из эпибласта и гипобласта, называется зародышевым щитком. Из зародышевого щитка образуется тело самого зародыша. Вторая фаза гаструляции – иммиграция связана со сложным перемещением клеток эпибласта сначала к заднему (хвостовому) концу зародыша (рис.13,А). Клетки эпибласта движутся двумя потоками по краям зародышевого диска, оба потока сталкиваются в хвостовом отделе у медианной (средней) линии, сливаются и затем направляются вперёд, к головному концу, образуя утолщенной клеточный тяж по средней линии – первичную полоску. Второй поток клеток (более медленный) распространяется со стороны головного конца зародыша и направляется по средней части эпибласта навстречу первому потоку. Место встречи двух потоков - первичный (гензеновский) узелок. Вдоль первичной полоски образуется углубление – первичная бороздка. Из первичной полоски клетки выселяются и вначале встраиваются в первичную энтодерму, раздвигая клетки гипобласта. Так формируется энтодерма зародыша. Далее клетки мигрируют из первичной полоски в обе стороны от неё, и встраиваются между первичной эктодермой и энтодермой, образуя средний зародышевый листок - мезодерму. В процессе гаструляции происходит также образование мезенхимы – эмбрионального зачатка соединительной ткани. Мезенхима выселяется из всех зародышевых листком, главным образом из мезодермы.
Рис.13. Иммиграция в эмбриогенезе птиц 1. первичный (гензеновский) узелок; 2. первичная ямка; 3. первичная полоска; 4. первичная бороздка; 5. эпибласт; 6. гипобласт; 7. клетки, выселяющиеся из первичной полоски; 8. мезодерма; 9. зародышевая энтодерма, замещающая гипобласт.
Из материала первичного узелка через первичную ямку мигрируют клетки, образующие клеточный тяж по средней линии зародыша – хордальный отросток (нотохорд). Из зародышевых листков и мезенхимы происходит формирование зачатков всех тканей (гистогенез) и органов (органогенез) развивающегося организма. Вначале закладываются осевые зачатки органов, которые располагаются по оси симметрии тела. Комплекс осевых органов включает: • нервную трубку • хорду • параксиальную мезодерму • кишечную трубку. Нервная трубка формируется из первичной эктодермы под влиянием сигнальных молекул, вырабатываемых клетками хорды, таким образом хорда является первичным индуктором нейроэктодермы. Процесс образования нервной трубки называется нейруляцией ( рис.14 ). В зоне образования нервной трубки в эктодерме вначале появляется утолщение – нервная пластинка. В нервной пластинке образуется углубление – нервный желобок, а по краям желобка утолщения – нервные складки. При смыкании нервных складок формируются нервная трубка и нервные валики (нервные гребни).
Рис.14. Схема нейруляции Из нервной трубки образуется центральная нервная система(головнойи спинной мозг). Клетки нервного гребня активно мигрируют и дают начало структурам периферической нервной системы и некоторым другим структурам (мозговое вещество надпочечников, эндокриноциты диффузной эндокринной системы, клетки-меланоциты и др. – табл.5). Та часть эпибласта, которая остаётся после выселения клеток из первичной полоски и нейруляции, образует зародышевую эктодерму (эпидермальную эктодерму ). Зародышевая эктодермаявляется источником развития эпителия кожного покрова и его производных. В мезодерме дифференцируются следующие части (рис.15): · сомиты (параксиальная мезодерма), которые располагаются справа и слева от нервной трубки и хорды; · нефротомы (сегментные ножки, промежуточная мезодерма), лежащие латеральнее сомитов; · спланхнотомы ( латеральная мезодерма), наиболее латерально расположенные участки мезодермы.
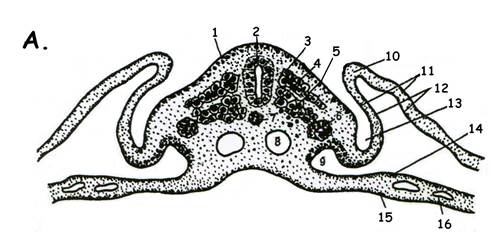
Рис.15. Зародыш курицы на этапе образования комплекса зачатков осевых органов. В.Сагиттальный срез 1.кожная эктодерма; 2.нервная трубка; 3.хорда; 4.мезодерма (сомит); 4.1. сомит (параксиальная мезодерма); 4.2.нефротом (промежуточная мезодерма, сегментные ножки); 4.3. спланхнотом (латеральная мезодерма); 4.3.1. – париетальный листок спланхнотома; 4.3.2. висцеральный листок мезодермы; 5.энтодерма; 6.целом;; 7.кровеносный сосуд.
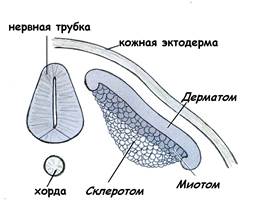
Сомиты и нефротомы представляют собой сегментированную часть мезодермы (рис 15А). В дальнейшем сомиты далее дифференцируются на:
· дерматом, из которого формируется соединительнотканная часть кожи – дерма; · миотом, из которого развивается скелетная мышечная ткань; · склеротом – источник развития тканей осевого скелета (рис.16).
Рис.16. Дифференцировка сомита Нефротом служит источником для развития мочевыделительной и половой систем. Спланхнотомы не сегментируются и расслаиваются на два листка – париетальный листок, прилежащий к эктодерме (соматическаямезодерма), и висцеральный листок, прилежащий к энтодерме (спланхническаямезодерма). Между париетальным и висцеральным листками формируется целомическая полость. Из листков спланхномезодермы происходит образование эпителия серозных оболочек, выстилающих брюшину, плевру, перикард.
Из мезенхимы развиваются: · кровь и лимфа; · сосуды; · кроветворные органы; · соединительная ткань; · гладкая мышечная ткань. Из энтодермы образуется эпителий среднего отдела кишечной трубки, крупных желез, связанных с пищеварительной трубкой (печень, поджелудочная железа), эпителий лёгкого, некоторых эндокринных желез (табл.4). ВНЕЗАРОДЫШЕВЫЕ (ПРОВИЗОРНЫЕ) ОРГАНЫ обеспечивают эмбриону условия для нормального развития и жизнедеятельности. Они функционируют в течение всего эмбриогенеза и исчезают по окончании эмбрионального развития, когда зародыш приобретает способность к самостоятельному существованию. Внезародышевые органы формируются из внезародышевых частей зародышевых листков. Внезародышевые органы у птиц: · амнион; · серозная оболочка; · желточный мешок; · аллантоис. Зародыш птиц первое время распластан из поверхности желтка, но в дальнейшем формируется туловищная складка. При этом все четыре листка (эктодерма, энтодерма, париетальный и висцеральный листки мезодермы) подгибаются под зародыш, отделяя его от внезародышевых органов (рис.17). Амниотические складки, образованные из внезародышевых частей эктодермы и париетального листка мезодермы, приподнимаются сначала над головным концом, а затем над всем телом зародыша (рис.17). Эктодерма одной амниотической складки срастается с эктодермой другой складки. Париетальные листки мезодермы обеих складок также срастаются в единый слой. При этом формируются заполненный жидкостью пузырь – амнион, стенка которого образована амниотической оболочкой, состоящей из эктодермы, выстилающей его изнутри, и париетального листка мезодермы, лежащего снаружи. Эктодермальный амниотический эпителий секретирует амниотическую жидкость, обеспечивая водную среду для развития зародыша и защищая его от механических повреждений. Желточный мешок образуется из внезародышевых частей энтодермы и висцерального листка мезодермы, которые постепенно обрастают поверхность желтка (рис.18). Желточный мешок отделяется от первичной кишки в процессе формирования туловищной складки. Желток переводится в растворимую форму под действием ферментов, которые продуцируются энтодермальными клетками (клетками желточной энтодермы), и используется зародышем для питания. По мере развития зародыша и потребления питательных веществ объём желточного мешка редуцируется. Серозная оболочка образована внезародышевыми частями париетального листка мезодермы (изнутри) и эктодермы (обращена наружу). Серозная оболочка обрастает весь зародыш и выполняет функцию дыхания (газообмена). Аллантоис возникает в виде полого выроста вентральной стенки задней части кишечной трубки и состоит из энтодермы зачатка задней кишки и покрывающего его снаружи висцерального листка мезодермы с сосудами. Аллантоис постепенно разрастается, занимая всё больший объём, приближается к серозной оболочке и снабжает её сосудами.
Рис.17. Дифференцировка зародышевых листков и формирование Внезародышевых органов А. 1.зародышевая эктодерма; 2.нервная трубка; 3.дерматом; 4.склеротом; 5.миотом; 6.нефротом; 7.хорда; 8.аорта; 9.целом; 10.амниотическая складка; 11.внезародышевая эктодерма; 12.париетальный листок спланхнотома; 13.туловищная складка; 14.висцеральный листок спланхнотома; 15.энтодерма; 16.кровеносные сосуды. В. 1.амнион; 2.серозная оболочка; 3.амниотическая складка; 4.туловищная складка; 5.целом; 6.желточный мешок; 7.аллантоис;
|
||||
|
|
Последнее изменение этой страницы: 2016-08-10; просмотров: 1183; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.1.23 (0.012 с.) |