
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Гладкая эндоплазматическая сеть.Содержание книги
Поиск на нашем сайте Краткий обзор: ЭПС- одномембранная органелла,представляет собой совокупность мембранных вакуолей, трубочек и плоских мешков (цистерн), распределённых тем или иным способом в цитоплазме. Основная часть: Гладкая (агранулярная) ЭПС - цистерн почти нет, с мембранами вакуолей и трубочек никакие видимые структуры не связаны. Функции: В мембраны гладкой ЭПС встроены ферменты гид- роксилирования (или микросомального окисления). Они необходимы - при синтезе многих липидов (например, стероидных гормонов) - и для обезвреживания токсических соединений. Гладкая ЭПС особенно выражена в клетках - надпочечников и гонад, синтезирующих стероидные гормоны, - и печени – в связи с синтезом холестерина.
Краткий обзор: Клеточный центр (центросома) — немембранный органоид, главный центр организации микротрубочек и регулятор хода клеточного цикла в клетках эукариот. Веретено́ деле́ния — динамичная структура, которая образуется в митозе и мейозе для обеспечения сегрегации хромосом и деления клетки. Полный ответ: Центриоли, обычно расположенные в паре (диплосома), и окруженные зоной более светлой цитоплазмы, от которой радиально отходят тонкие фибриллы (центросфера). Совокупность центриолей и центросферы называют клеточным центром. Совокупность центриолей и центросомы называют клеточным центром.
В интерфазе клеточного цикла центросомы ассоциированы с ядерной мембраной. В профазе митоза ядерная мембрана разрушается, центросома делится, и продукты её деления (дочерние центросомы) мигрируют к полюсам делящегося ядра. Микротрубочки, растущие из дочерних центросом, крепятся другим концом к так называемым кинетохорам на центромерах хромосом, формируя веретено деления. По завершении деления в каждой из дочерних клеток оказывается только по одной центросоме. Помимо участия в делении ядра, центросома играет важную роль в формировании жгутиков и ресничек. Центриоли, расположенные в ней, выполняют функцию центров организации для микротрубочек аксонем жгутиков. У организмов, лишенных центриолей (например, у сумчатых), жгутики не развиваются. Веретено́ деле́ния — динамичная структура, которая образуется в митозе и мейозе для обеспечения сегрегации хромосом и деления клетки. Веретено образуют три основных структурных элемента: микротрубочки, полюса деления и хромосомы. В организации полюсов деления у животных участвуют центросомы, содержащие центриоли. Важную роль в формировании веретена играют моторные белки, относящиеся к семействам динеинов и кинезинов. Полноценное веретено деления образуется на стадии прометафазы после разрушения ядерной мембраны. Веретено деления типичной клетки млекопитающих состоит из трёх структурных элементов — центросом, микротрубочек и хромосом, — которые образуют симметричную биполярную структуру. На полюсах веретена располагаются центросомы — небольшие органеллы, функционирующие как центры организации микротрубочек. Каждая центросома состоит из пары центриолей, окруженных множеством разных белков. Между полюсами веретена находятся конденсированные хромосомы, состоящие из пары хроматид, скреплённых в области центромеры. На цетромерных участках хромосом находятся кинетохоры — сложные структуры, отвечающие за прикрепление к микротрубочкам веретена[1]. Веретено деления состоит из двух полуверетён. Полуверетено образуется из поляризованных микротрубочек. Отрицательные минус-концы микротрубочек собираются на полюсах веретена вокруг центросом. Плюс-концы микротрубочек отдаляются от двух полюсов и пересекаются в средней экваториальной части веретена. У большинства позвоночных полуверетено состоит из 600—750 микротрубочек, 30—40 % которых заканчиваются на кинетохорах. Микротрубочки, которые соединяют полюса веретена с кинетохорами хромосом, называются кинетохорными. Причём каждый кинетохор при образовании веретена связывается с множеством микротрубочек и образует кинетохорный пучок. Микротрубочки, которые располагаются между полюсами и не присоединяются к кинетохорам, называются межполюсными. Часть микротрубочек веретена образует вокруг каждого полюса радиальные структуры, называемые звёздами или астерами. Такие микротрубочки называются астральными. Вопрос №11 Митохондрии Краткий обзор (из методички) – Двумембранные образование, имеющие собственную ДНК, предположительно возникли из прокариот после объединение с эукариотическими клетками в результате эволюции и последующего с ними сосуществования (симбиоза). Обеспечивают синтез АТФ за счет реакций окислительного фосфорилирования. Митохондрии контролируют внутриклеточное содержание ионов кальция, обеспечивают образование тепла, а также запрограммированной гибели клеток. Основная часть (из инета) - Это органеллы размером с бактерию. Митохондрия ограничена двумя мембранами - гладкой внешней и складчатой внутренней, имеющей очень большую поверхность. Складки внутренней мембраны глубоко входят в матрикс митохондрий, образуя поперечный перегородки - кристы. Пространство между внешней и внутренней мембранами обычно называют межмембранным пространством. Мембраны митохондрий содержат интегральные мембранные белки. Во внешнюю мембрану входят порины, которые образуют поры и делают мембраны проницаемыми для веществ с молекулярной массой до 10 кДа. Внутренняя же мембрана митохондрий непроницаема для большинства молекул; исключение составляют О2, СО2, Н20. Внутренняя мембрана митохондрий характеризуется необычно высоким содержанием белков (75%).В их число входят транспортные белки-переносчики, ферменты, компоненты дыхательной цепи и АТФ-синтаза. Кроме того, в ней содержится необычный фосфолипид кардиолипин. Матрикс также обогащен белками, особенно ферментами цитратного цикла. Метаболические функции Митохондрии являются «силовой станцией» клетки, поскольку за счет окислительной деградации питательных веществ в них синтезируется большая часть необходимого клетке АТФ (АТР). В митохондриях локализованы следующие метаболические процессы: превращение пирувата в ацетил-КоА, катализируемое пируватдегидрогеназным комплексом: цитратный цикл; дыхательная цепь, сопряженная с синтезом АТФ (сочетание этих процессов носит название «окислительное фосфорилирование»); расщепление жирных кислот путем β-окисления и частично цикл мочевины. Митохондрии также поставляют клетке продукты промежуточного метаболизма и действуют наряду с ЭР как депо ионов кальция, которое с помощью ионных насосов поддерживает концентрацию Са2+ в цитоплазме на постоянном низком уровне (ниже 1 мкмоль/л). Главной функцией митохондрий является захват богатых энергией субстратов (жирные кислоты, пируват, углеродный скелет аминокислот) из цитоплазмы и их окислительное расщепление с образованием СО2 и Н2О, сопряженное с синтезом АТФ. 12. Рибосомы. Полирибосомы. Митохондриальные рибосомы. Краткий обзор: Рибосома- немембранный двухсубъкдиничный (малая и большая субъединицы) органоид, состоящий из рРНК и белков. Рибосомы имеют сферическую или слегка эллипсоидную форму. Полисома, или полирибосома — несколько рибосом, одновременно транслирующих одну молекулу мРНК. Рибосомы митохондрий, или миторибосомы, ассоциированы с митохондриальным матриксом. Основная часть: Рибосома- немембранный двухсубъкдиничный (малая и большая субъединицы) органоид, состоящий из рРНК и белков. Рибосомы имеют сферическую или слегка эллипсоидную форму. Служит для биосинтеза белка из аминокислот. Рибосомы подразделяются на свободные и связанные с мембранами ЭПС и наружной ядерной мембраной. Свободные рибосомы синтезируют синтезируют белки для самой клетки, а связанные- на экспорт. Синтез рибосом у эукариот происходит в специальной внутриядерной структуре —ядрышке. Полисома, или полирибосома — несколько рибосом, одновременно транслирующих одну молекулу мРНК. Рибосомы митохондрий, или миторибосомы, ассоциированы с митохондриальным матриксом. Вопрос №13 Аппарат Гольджи Краткий обзор. Аппарат Гольджи — это система внутриклеточных мембранных структур: цистерн и пузырьков, в которых накапливаются вещества, синтезированные на мембранах ЭПС. Вещества доставляются в комплекс Гольджи в мембранных пузырьках, которые отшнуровываются от эндоплазматической сети и присоединяются к цистернам комплекса Гольджи. Здесь эти вещества претерпевают различные биохимические превращения, а затем снова упаковываются в мембранные пузырьки, и большая их часть транспортируется к цитоплазматической мембране. Мембрана пузырьков сливается с цитоплазматической мембраной, а содержимое выводится за пределы клетки посредством экзоцитоза. В комплексе Гольджи растительных клеток синтезируются полисахариды клеточной стенки (оболочки). Еще одна важная функция комплекса Гольджи — это образование лизосом. Комплекс Гольджи был открыт в 1898 г. итальянским гистологом Камилло Гольджи В нервных клетках. Основная часть. Комплекс Гольджи — это место конденсации и накопления продуктов секреции, вырабатываемых в других участках клетки, в основном в ЭПС. На наружной, вогнутой стороне стопки из пузырьков постоянно формируются новые цистерны, а на внутренней стороне цистерны превращаются обратно в пузырьки. При световой микроскопии он может распределяться в виде сложных сетей или отдельных диффузно расположенных участков (диктиосом). Форма и положение органеллы не имеют принципиального значения и могут изменяться в зависимости от функционального состояния клетки. При электронной микроскопии видно, что комплекс состоит из скоплений плоских цистерн В целом комплекс Гольджи участвует в сегрегации — это разделение, отделение определенных частей от основной массы, и накоплении продуктов, синтезированных в ЭПС, в их химических перестройках, созревании. Секреторная функция комплекса Гольджи заключается в том, что синтезированный на рибосомах экспортируемый белок, отделяющийся и накапливающийся внутри цистерн ЭПС, транспортируется в вакуоли пластинчатого аппарата. Комплекс Гольджи может резко увеличиваться в размерах в клетках, активно осуществляющих секреторную функцию,обычно сопровождается развитием ЭПС, а в случае синтеза белков — ядрышка. Во время деления клетки комплекс Гольджи распадается до отдельных цистерн или пузырьков, которые распределяются между двумя делящимися клетками и в конце телофазы восстанавливают структурную целостность органеллы. Вне деления происходит непрерывное обновление мембранного аппарата за счет пузырьков, мигрирующих из ЭПС и дистальных цистерн диктиосомы за счет проксимальных компартментов. о содержимого в процессе лизосомного экзоцитоза. 14. Лизосомы- одномембранные структуры, образуются путем слияния перинуклеарных эндосом,содержащих лизосомные гидролазы и лизосомные мембранные белки, с везикулами, подлежащими деградации (периферической эндосомой, фагосомой или аутофагоцитозной вакуолью). * перинуклеарные эндосомы образуются при слиянии везикул, содержащие лизосомные гидролазы после их синтеза в гранулярной эпс и процессинга в комплексе Гольджи, и везикул, в мембрану которых встроены специфические лизосомные мембранные белки. * периферические эндосомы образуются в результате эндоцитоза. * мультивезикулярные тельца образуются при слиянии перинуклеарной и периферической эндосом. * фаголизосома образуется при слиянии перинуклеарной эндосомы и фагосомы. * аутофаголизосома образуется при слиянии перинуклеарной эндосомы и аутофагоцитозной вакуоли, содержащей подлежащие деградации эндогенные молекулы и органеллы. * остаточные тельца - лизосомы любого типа, содержащие непереваренный материал (липофусцин, гемосидерин). Функция: Катализ гидролитического (в водной среде) расщепления нуклеиновых кислот, белков, жиров, полисахаридов и мукополисахаридов, других химических соединений при низких значениях pH. В цитобиологических понятиях - внутриклеточное переваривание веществ и структур. Пероксисомы - одномембранные органеллы, пузырьки с электроноплотной сердцевиной. В составе мембраны органеллы находятся специфичные белки - пероксины, а в матриксе - более 40 ферментов, каиализирующих анаболичиские (биосинтез желчных кислот, H2O3, зависимое дыхание, деградация ксенобиотиков) процессы.
|
||||
|
|
Последнее изменение этой страницы: 2016-08-14; просмотров: 696; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.170 (0.008 с.) |


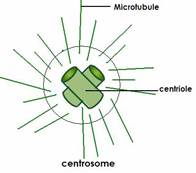 10. Центросома. Веретено деления. Молекулярное строение и функция.
10. Центросома. Веретено деления. Молекулярное строение и функция. Клеточный центр (центросома) — немембранный органоид, главный центр организации микротрубочек и регулятор хода клеточного цикла в клетках эукариот. Важная роль его заключается в клеточном делении, в частности, в образовании митотического веретена. Впервые обнаружена в 1883 году Теодором Бовери, который назвал её «особым органом клеточного деления. В клетке содержится одна или две центросомы.
Клеточный центр (центросома) — немембранный органоид, главный центр организации микротрубочек и регулятор хода клеточного цикла в клетках эукариот. Важная роль его заключается в клеточном делении, в частности, в образовании митотического веретена. Впервые обнаружена в 1883 году Теодором Бовери, который назвал её «особым органом клеточного деления. В клетке содержится одна или две центросомы. Каждая центриоль построена из 27 цилиндрических элементов (микротрубочек), сгруппированных в 9 триплетов образованных в результате полимеризации белка тубулина. Девять триплетов микротрубочек расположены по окружности, образуя таким образом полый цилиндр.
Каждая центриоль построена из 27 цилиндрических элементов (микротрубочек), сгруппированных в 9 триплетов образованных в результате полимеризации белка тубулина. Девять триплетов микротрубочек расположены по окружности, образуя таким образом полый цилиндр. Типичное веретено является биполярным — между двумя полюсами образуется веретенообразная система микротрубочек. Микротрубочки веретена присоединяются к кинетохорам хроматид в области центромер и обеспечивают движение хромосом по направлению к полюсам.
Типичное веретено является биполярным — между двумя полюсами образуется веретенообразная система микротрубочек. Микротрубочки веретена присоединяются к кинетохорам хроматид в области центромер и обеспечивают движение хромосом по направлению к полюсам.


