Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Аннотирование геномов. Белок-кодирующие последовательности и их предсказания.Содержание книги
Поиск на нашем сайте
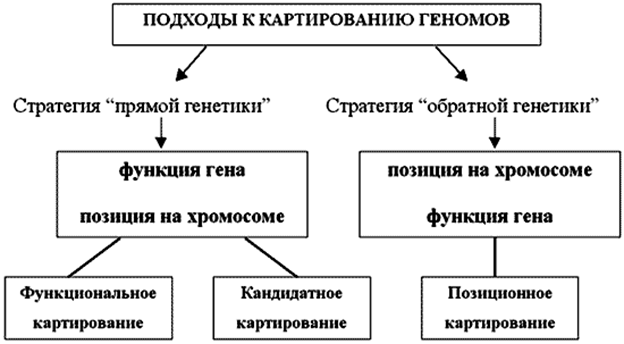
Аннотация генома: описание функциональных (тканеснецифичность экспрессии генов-профили экспрессии, взаимосвязи между генами) и структурных (местонахождение кодирующих участков генов, регуляторных элементов) характеристик генома. Основной метод, как всегда, ПЦР (аннотирование невозможно без картирования, а физическое и цитогенетическое картирование – без секвенирования). Часто используют библиотеки кДНК (в т.ч. для генов, кодирующих структурные белки). Сравнивают сходство и различие АК и н.п. последовательностей, если схожи н.п. и тем более АК –предполагают сходство функции белка; если возможно, проверяют, синтезируя белки по АК и изучая их свойства. АК-предпочтительнее (эволюционируют медленнее, устойчивее к мутациям - замена 3-го н. в кодоне (синонимичные замены) обычно не влияет на АК; обычно короче, чем н.п.), из н.п.- чаще мРНК (старт и стоп-кодон-хорошие индикаторы, но они встречаются и в не-белок кодирующих н.п.). О возможности правильного сплайсинга можно судить по н.п. интронов (начало - GT,конец -AG), но не всегда достоверно. Нужно помнить о вырожденности генетического кода, различной предпочтительности кодонов и отклонениях от стандартного генетического кода, в зависимости от изучаемого генома. Разные н.п. могут кодировать схожие белки, поскольку АК одни и те же (вырожденность). У ПРО частота использования кодона прямо пропорциональна концентрации типов тРНК – только та, С которой максимальна, используется для синтеза белка; содержание GC-пар в геноме схоже с содержанием GC-пар только в 3-й позиции кодона (действует только мутационное давление, зависящее от скорости прямых и обратных мутаций, их синонимичности), во второй действует элиминирующий отбор (дефектный белок-гибель организма), в первой-оба; у ЭУ, особенно позвоночных, такнеспецифичность привела к выраженной мозаичности генома – GC-богатые(60%) и GC-бедные (30%) участки хр. Примеры. Аргинин у E.coli кодируется GGU и GGC, не используются CGA,CGG,AGA,AGG, хотя тоже кодируют аргинин. В мт-геноме UGA кодирует триптофан и не является терминирующим, AGA,AGG-терминирующие, а не кодируют аргинин, AUA кодирует метионин вместо изолейцина, в мит. растений CGG редактируется в UGG, прежде чем кодировать триптофан; UAA,UAG у простейших Tetrahymena,Paramecium в ядерных генах кодируют глутамин (а не стоп-кодон); у Mycoplasma capricolum UGA кодирует триптофан. Созданы сложные математические модели для расчёта как эволюционных расстояний АК-последовательностей, так и для определения вероятности замен (схожие модели созданы для подсчёта количества нуклеотидных замен и их вероятности, однако даже эти показатели видо-и иногда ткане-специфичны). В случае изучения н.п., преимущественно используется физическое картирование (маркерами служат короткие повторы, сайты рестрикции, минисателлиты, микросателлиты, фрагменты амплификации с произвольными либо «конкретными» праймерами, snp, ретротранспозоны, экспрессирующиеся н.п.; маркеры должны обладать полиморфизмом) и цитогенетические методы (основаны на гибридизации н.п., позволяет предположить «родственные связи» и время дивергенции как организмов, так и н.п., кодирующих интересующий белок). Существует 4 подхода локализации гена. Функциональное картирование: выделение в чистом виде белкового продукта гена, далее к нему по аминокислотной последовательности подбирают вырожденные праймеры и проводят ПЦР-скрининг геномных библиотек. Однако список генов, для которых эта информация была достаточно полной к настоящему времени практически исчерпан и большинство генов, функция которых была известна, уже клонированы и локализованы. Появление в конце 80-х годов множества высокополиморфных ДНК-маркеров дало возможность пойти в обратном направлении - от хромосомной карты к функции. Место гена (функция неизвестна) на карте устанавливают по результатам анализа сцепления гена с ранее локализованными генетическими маркерами и далее детально исследуется уже область генома рядом с маркером. Главным ограничением позиционного подхода является низкая разрешающая способность генетических карт - интервал между двумя соседними маркерами, в котором локализован ген, может оказаться слишком велик и недоступен физическому картированию. Для большинства генов, которые были локализованы, характерны структурные аномалии (как правило, это гены, ответственные за наследственные заболевания человека), что существенно облегчает заключительную стадию поиска гена - выделение и локализацию гена. Сейчас используют позиционно-кандидатное картирование - поиск на выявленном участке генома подходящих кандидатных генов (предполагается, что они кодируют нужный белок; можно синтезировать белок и проверить, или использовать мультиплексную(с несколькими праймерами) либо вложенную (2 пары праймеров, 2 ПЦР – продукты первой служат матрицей для второй) ПЦР).
|
||||
|
|
Последнее изменение этой страницы: 2016-07-11; просмотров: 378; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 13.59.1.58 (0.007 с.) |