Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Башкирский государственный педагогическийСодержание книги
Поиск на нашем сайте МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ ГОСУДАРСТВЕННОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ БАШКИРСКИЙ ГОСУДАРСТВЕННЫЙ ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ им. М.АКМУЛЛЫ
БИОЛОГИЯ РАЗМНОЖЕНИЯ И РАЗВИТИЯ
КУРС ЛЕКЦИЙ
УФА 2006 УДК 576.4 ББК 28.073 Б 63 Печатается по решению редакционно-издательского совета Башкирского государственного педагогического университета им. М.Акмуллы Биология размножения и развития: курс лекций [Текст] / сост. О.А. Абросимова; под ред. В.Ю. Горбуновой. – Уфа: Издательство БГПУ, 2006. – 140с.
Лекционный курс разработан в соответствии с Государственным образовательным стандартом и учебной программой курса «Биология размножения и развития» для студентов, обучающихся по специальности 012100 «Генетика». Материал излагается в соответствии со стадиями развития организмов – от прогенеза до процесса органогенеза. Рассматриваются особенности развития анамний и амниот, а также эмбриогенез и проблема биологического возраста человека. Включен материал о генетической и гормональной регуляции процесса индивидуального развития организмов, о процессах регенерации. Рассмотрены вопросы становления биологии индивидуального развития и ее фундаментальные и прикладные задачи. Может быть полезен для студентов университетов, обучающихся по биологическим специальностям.
Автор–составитель: О.А. Абросимова, канд. биол. н., доц.
Научный редактор: В.Ю. Горбунова, д-р биол. н., проф.
Рецензенты: В.Г. Шамратова, д-р биол. н., проф. (БГУ); Р.Р. Кабиров, д-р биол. н., проф. (БГПУ).
© Издательство БГПУ, 2006
ПРЕДИСЛОВИЕ
Биология развития является наукой, охватывающей наибольший спектр биологических проблем. Она играет объединяющую роль в отношении разных биологических наук и создает основу для интеграции молекулярной биологии, физиологии, биологии клетки, анатомии, онкологии, иммунологии и даже эволюционных и экологических исследований. Изучение эмбрионального развития организмов стало необходимым для понимания любой другой области биологии. Многоклеточные организмы формируются в результате сравнительно медленного процесса прогрессивных изменений, которые называются развитием. Обычно развитие многоклеточного организма начинается с одной клетки – оплодотворенного яйца, или зиготы, которая митотически делится и дает начало всем клеткам организмов. Наука, изучающая развитие животных, по традиции называется эмбриологией, т.к. развивающийся организм на стадиях от оплодотворенного яйца до рождения носит название эмбриона или зародыша. Однако развитие не останавливается не только в момент рождения, но и даже по достижении взрослого состояния. У большинства организмов развитие не прекращается на протяжении всей их жизни. Например, у человека каждый день происходит замена более 1 грамма клеток кожи, поскольку старые клетки слущиваются, и ежеминутно в течение жизни в костном мозге образуются миллионы новых эритроцитов. Поэтому о биологии развития принято говорить как о науке, включающей изучение не только эмбриогенеза, но и других процессов развития. Исследования в любой области биологии стали невозможными без знания основ биологии развития. В предлагаемом учебном пособии материал расположен в определенной последовательности, в которой предшествующие лекции могут служить основой для понимания последующих. Это облегчает изучение материала каждой новой лекции. ЛЕКЦИЯ 1
ИСТОКИ И ОСНОВНЫЕ ЭТАПЫ СТАНОВЛЕНИЯ БИОЛОГИИ ИНДИВИДУАЛЬНОГО РАЗВИТИЯ ПЛАН Введение. 1. Представления о развитии организмов в трудах мыслителей древности. 2. Развитие эмбриологии в XVI – XVII веках. 3. Вклад К.Ф.Вольфа и К.Бэра в дальнейшее развитие эмбриологии. 4. Формирование и развитие аналитической и экспериментальной эмбриологии. 5. Участие советских ученых в дальнейшем развитии эмбриологии. 6. Исследования ученых-генетиков в области эмбриологии. 7. Начало работ в области биохимической генетики. 8. Становление биологии индивидуального развития.
Введение Биология индивидуального развития – область науки, изучающая закономерности онтогенетического развития организмов. Она сформировалась в последние десятилетия на основе достижений экспериментальной эмбриологии, молекулярной биологии, генетики, цитологии. Задача биологии индивидуального развития – исследование макро- и микроморфологических, физиолого-биохимических, молекулярных и генетических процессов, протекающих в развивающейся особи, выяснение факторов и механизмов, управляющих процессами развития на всех этапах онтогенеза животных, растительных организмов, а также одноклеточных форм. Столь широкий охват обусловлен распространенностью элементарных и общих закономерностей процессов развития в живой природе. Начало разработки проблем, которыми сегодня занимается биология индивидуального развития, восходит к 70–80-м годам прошлого столетия, когда на основе успехов сравнительной и эволюционной эмбриологии зародились методология и основные тенденции аналитической и экспериментальной эмбриологии, сформировались первые концепции о цитоэмбриологических механизмах наследственности. В начале века был создан фундамент всех тех областей эмбриологии, цитологии, генетики, биохимии, которые в последующем легли в основу биологии индивидуального развития. Бурный прогресс молекулярной биологии создал условия для дальнейшего их объединения, открыв доступ к пониманию наиболее тонких молекулярных механизмов эмбрио-цито-генетических аспектов процесса развития. В настоящее время знания в этой области продолжают углубляться и расширяться. Это создает трудности в однозначном и четком определении целей, методов и объектов биологии индивидуального развития. Их преодоление связано с дальнейшей разработкой проблем, относящихся к этой области, с учетом особенностей индивидуального развития организмов, отличающихся уровнем организации. Для рассматриваемой области биологии характерен синтетический подход к изучению процессов развития, так как ее цель – познание закономерностей этих процессов применительно к целостнойживой системе, создание единой концепции онтогенеза и разработка способов управления им. Управление онтогенезом – необходимое условие для решения многих актуальных задач медицины и сельского хозяйства, прежде всего животноводства. Биология индивидуального развития животных призвана внести весомый вклад в реализацию задач Продовольственной программы по резкой интенсификации работ, направленных на выведение новых, ценных для животноводства пород сельскохозяйственных животных и их ускоренное воспроизводство. В последние годы в этой области биологии созданы фундаментальные и технические предпосылки для решения таких сложных и актуальных народнохозяйственных задач. На основе достижений экспериментальной эмбриологии, а также клеточной и генной инженерии разрабатываются методы манипулирования с яйцеклетками, эмбрионами сельскохозяйственных животных и их наследственным аппаратом с целью направленного вмешательства в процессы их воспроизводства и наследственность. Успешная реализация этих перспективных возможностей требует подготовки специалистов, способных проводить исследования на стыке эмбриологии, генетики, цитологии, молекулярной биологии.
Мыслителей древности
Биология индивидуального развития – это область, формирующаяся в последние десятилетия путем синтеза ряда биологических дисциплин – эмбриологии, генетики, цитологии, биохимии и физико-химических направлений биологии (молекулярная биология, биоорганическая химия, биофизика). Процесс синтеза этих достижений в рамках биологии развития еще продолжается, и пока существует много неясного в вопросах о роли и вкладе той или иной дисциплины, их взаимоотношениях. Возникает, в частности, вопрос, имеется ли у такой широкой по профилю, многоплановой области науки единая методология и единство в понимании целей и задач, необходимых для любой научной дисциплины. Дискуссии по поводу этих вопросов будут продолжаться до тех пор, пока в рамках этой области продолжается объединение методов и знаний разнообразных дисциплин вокруг общей задачи – познания фундаментальных принципов и механизмов индивидуального развития живых существ всех уровней организации и разработка на этой основе методов управления воспроизводством и онтогенетическим развитием. Как сориентироваться в современной биологии индивидуального развития, в ее методологии, как определить будущее этой науки, выделить главные ее тенденции? Понять будущее науки помогает анализ прошлого, в данном случае – история становления биологии индивидуального развития, выявление ее истоков, роли и вклада в нее отдельных дисциплин. Методология, основные цели и задачи биологии индивидуального развития на протяжении длительного времени формировались главным образом в области эмбриологии с ее многовековой историей борьбы, которую вели сторонники прогрессивной научной мысли. В современную эпоху эмбриология также остается основой биологии развития, потому что ей в наибольшей мере свойственны синтетический подход, понимание значения категорий целостности в индивидуальном развитии. Таким образом, эмбриология, вложив в биологию индивидуального развития основные методологические принципы и объединяя вокруг себя аналитические науки, направляет их достижения на решение главной задачи – раскрытие закономерностей индивидуального развития целостной живой системы. Остановимся на некоторых этапах истории эмбриологии. Эмбриология – одна из наиболее древних наук. На протяжении длительной истории эта наука, олицетворяя глубокий интерес человека к тайнам зарождения живых существ, впитывала научно-технические и идейно-философские достижения, способствовала формированию научной идеологии. Вопросы зарождения и развития новых особей были предметом многочисленных религиозных легенд и мифов. Не останавливаясь на этих моментах предыстории, а также на сведениях о примитивных эмбриологических представлениях в Египте и Древнем Востоке, обратимся к Древней Греции – стране, где впервые в истории человечества началась осознанная работа над теоретико-философскими основами наук. Древнегреческому врачу и философу Гиппократу (460–377 г. до н. э.) приписывают сборник, в котором содержатся первые научные высказывания о развитии эмбрионов и факторах формообразования. В своих воззрениях Гиппократ основывался на предположении, что зародыш строится под действием «внутреннего огня»: части, более податливые огню, выгорают и на их месте образуются полости, другие – лишь ссыхаются и уплотняются и из них получаются стенки полостей. В результате возникают, например, органы пищеварительного тракта. По мнению Гиппократа, явления органического развития вполне можно объяснить свойствами неорганической материи. Кроме того, в их основе заложены принципы, которые позже стали характерны для концепций преформизма. Отличительная черта преформационных концепций – признание существования изначальных различий между частями зародыша (у Гиппократа – различия в «податливости» к огню) и мнение, что «отделение частей» (дифференцировка) происходит лишь в некоторый начальный момент развития, а в дальнейшем разделившиеся части только растут. Во многом своеобразными были взгляды одного из величайших мыслителей древности, основоположника естественных наук Аристотеля (384–322 до н. э.). Эмбриологические факты, которыми располагал Аристотель, и его воззрения на развитие жизни подробно изложены в сочинении «О возникновении животных». Аристотель знал о развитии куриного зародыша уже почти все, что можно увидеть без специальной обработки и невооруженным глазом. Он имел немалые сведения по анатомии и физиологии организмов. Однако огромное влияние Аристотеля на последующую науку связано не столько с его фактическими наблюдениями, сколько с теоретико-философскими представлениями. Аристотель счел недостаточными для объяснения развития те, по современной терминологии, «механистические» причины, к которым сводил развитие Гиппократ. По Аристотелю, все природные явления определяются не только наличием нужного «неоформленного» материала («материальная причина») и начальным толчком («действующая причина»), но также «формальной (финальной) причиной» развития. Последняя есть «цель» данного процесса, та форма, к которой этот процесс стремится. В противоположность Гиппократу Аристотель считал (и аргументировал конкретными примерами), что органы возникают не все сразу, а постепенно, один вслед за другим из бесструктурной вначале массы. Такое представление сделало Аристотеля основателем эпигенеза – противоположного преформизму учения о постепенном развитии, связанном с усложнением организации.
Развитие эмбриологии Решительный поворот в эмбриологии был осуществлен в 1759 г. петербургским академиком Каспаром Фридрихом Вольфом (1734–1794). В этом году Вольф представил свою диссертацию под названием «Теория зарождения». В те времена господствовало мнение физиолога и анатома А.Галлера о том, что трубчатые и мешкоподобные структуры зародыша (например, его кишечник) с самого начала имеют такую же форму, но это трудно заметить из-за тонкости стенок и их плотного слипания. Позже происходит их простое раздувание. Такое толкование строго соответствовало преформационной теории. Вольф установил совершенно иное. Кишечник, а также зачаток нервной системы сначала представляют собой пласты, которые лишь позже скручиваются в трубки. В ходе развития образуются новые формы. По сути дела Вольф открыл формообразование и тем самым дал первый позитивный и неопровержимый аргумент в пользу эпигенеза. Судьба этих, казалось бы, столь ясных работ была трудной. Под давлением господствующих авторитетов выводы Вольфа отвергались, и его работы были на некоторое время забыты. Надо сказать, что еще при жизни Вольфа с весьма остроумными доводами в пользу эпигенеза выступил немецкий профессор И.Ф.Блюменбах (1752–1840). Он впервые указал на несовместимость с преформизмом всевозможных случайных новообразований (например, галлы у растений) или регенерации гидры из любого, произвольно выбранного участка тела. О большой его наблюдательности и прозорливости свидетельствует то, что он обнаружил регуляции формы организма, не связанные с его ростом. Так, целая гидра восстанавливается из своей продольной половинки простым схождением краев разреза, тогда как, по убеждениям преформистов, такой процесс должен быть обязательно связан с ростом. Таким образом, для Блюменбаха, как и для Вольфа, одним из основных аргументов против преформизма было обнаружение «чистого», не связанного с ростом формообразования. Но несмотря на эти единичные догадки, уровень естественных наук не позволял еще обрести им прочную основу. Дальнейший прогресс в эмбриологии позвоночных связан с именами М.Ратке (1793–1860), X.Пандера (1794–1865) и К.Бэра (1792–1876). Пандер в 1817г. впервые описал зародышевые листки. Он нашел, что зародыш цыпленка на определенной стадии состоит из трех пластов: наружного – серозного, самого глубокого – слизистого и промежуточного – кровяного. К.Бэр распространил этот структурный принцип на всех позвоночных, обнаружив такие же листки в развитии рыб, лягушки и черепахи. Однако у зародышей птиц он насчитывал четыре листка, считая за отдельный листок каждый из двух слоев, на которые в ходе развития расслаивается мезодерма. Таким образом, Бэр установил единство плана строения зародышей разных классов позвоночных. Это привело его к важнейшему обобщению – «закону зародышевого сходства». Бэр утверждал, что зародыши разных видов, относящихся к одному типу, более сходны между собой, нежели взрослые формы, и что их видовые различия в ходе развития постепенно нарастают. Иными словами, сначала в развитии проявляются черты типа, потом класса и т. д. Бэр – автор многих важнейших открытий. Он впервые правильно описал яйцо млекопитающих и человека (1827) и хорду зародышей позвоночных. В споре преформистов с эпигенетиками Бэр занимал осторожную промежуточную позицию. Всецело соглашаясь с фактическими выводами Вольфа, он выступал против утверждений о полной «бесструктурности» ранних закладок. Бэр подчеркивал преемственность каждого этапа развития – от более простого к более сложному. По его словам, развитие есть не предобразование, не новообразование, а преобразование. Такая точка зрения полностью подтверждена последующим ходом развития науки. Следующий важнейший идейный перелом в эмбриологии, как и вообще в биологии, связан с выходом в свет в 1859 г. «Происхождения видов» Ч.Дарвина. Дарвинизм прежде всего подрывал главную опору телеологического мировоззрения, указывая на относительность органической целесообразности и на возможность достижения ее методом «проб и ошибок» (теория естественного отбора). Именно это произвело наибольшее впечатление на современников. Но не только этим своим аспектом дарвинизм повлиял на развитие эмбриологии. Наряду с палеонтологией и сравнительной анатомией Дарвин обращался к эмбриологии в поисках подтверждения своей эволюционной теории. По его словам, «...в высшей степени вероятно, что зародышевые или личиночные стадии многих животных более или менее ясно указывают на строение прародителя всей группы в его взрослом состоянии». Таким образом, Дарвин предлагал эволюционное истолкование закона Бэра. В более категоричной форме это же положение было выражено в биогенетическом законе Э.Геккеля (1834–1919): «онтогенез есть краткое повторение филогенеза». Гипотеза Дарвина оказалась мощным стимулом к эмбриологическим исследованиям. На основе эволюционной теории ученые разных стран за считанные годы выяснили развитие обширных, ранее совершенно не изученных групп организмов. Среди них первыми были русские эмбриологи А.О.Ковалевский (1840–1901) и И.И.Мечников (1845–1916). Особое значение имели работы Ковалевского по развитию ланцетника и асцидий, в которых были продемонстрированы сходные черты в развитии позвоночных и беспозвоночных животных. Эти исследования способствовали укреплению взглядов на эволюцию как на сквозной монофилетический процесс. А.О.Ковалевский – один из основоположников теории зародышевых листков. Развитии эмбриологии В разработке такого понимания процессов развития, при котором одна и та же система факторов развития зародыша определяет и ее целостность, и ее дифференциацию на части, важная заслуга принадлежит советскому биологу А.Г.Гурвичу (1874–1954). Он искал такие факторы, действием которых можно было объяснить интегрированность процессов развития, их пространственную организацию. Гурвич начинал свою научную деятельность под сильным влиянием идей Дриша, но не разделял его взглядов на непознаваемость целостных факторов развития. Он описывал эти факторы и их отношения с частями организма в точных математических выражениях, что для того времени (начало XX в.) было новым и непривычным. Гурвич первый ввел статистические методы в эмбриологию и обнаружил явление "нормировки" клеточных делений в целом организме, а также участие случайных событий в развитии. Ему принадлежат первые математические модели развивающихся систем, в ряде отношений предвосхитившие современные. Своей основной целью А.Г.Гурвич считал построение теории биологического поля, которую он постоянно видоизменял и совершенствовал, стремясь найти все более точные и близкие к физике формулировки взаимодействия частей в развивающемся организме. Новый этап в изучении механизмов развития связан с исследованиями школы М.М.Завадовского, создавшего направление, названное им динамикой развития. Завадовский придавал большое значение физиологическим аспектам развития, гуморальной среде, гормонам и т.д. Постепенно формировалось направление «химической эмбриологии» (Дж.Нидхем), интенсивно развивались цитология и генетика индивидуального развития. Все это открывало новые возможности для более детального изучения различных сторон индивидуального развития, для углубления в детали процессов. Вместе с тем основные принципы и категории науки об индивидуальном развитии, ее методология, сформированные в аналитической и экспериментальной эмбриологии, остались непоколебимыми, определяя и в дальнейшем главные направления поисков. Это не означает, что принципы и категории развития, сформированные в рамках экспериментальной эмбриологии, не испытали сильного влияния со стороны других дисциплин. Напротив, такое влияние было значительным, и без его рассмотрения нельзя понять истинную историю и логику становления современной биологии индивидуального развития. Одно из важных направлений эмбриологии, возникшее в СССР, связано с именем Д.П.Филатова (1876–1943). Он обосновал сравнительно-морфологический подход в экспериментальной эмбриологии, который был направлен на устранение накопившихся к этому времени противоречий между сравнительно-эволюционной и экспериментальной эмбриологией. Он ввел представление о «формообразовательном аппарате» как системе двусторонних взаимодействий между индуктором и реагирующей тканью, первым отметил неспецифичность ранних этапов детерминационного процесса, обосновал (одновременно с Г.Шпеманом и Ф.Леманом) принцип комплексности развития. П.Филатов создал крупную школу советских эмбриологов–экспериментаторов, из которых многие (В.В.Попов, Т.А. Детлаф и др.) внесли существенный вклад в науку. Среди выдающихся советских эмбриологов следует отметить П.П.Иванова (1878–1942) – автора теории о ларвальном и постларвальном отделах тела первичноротых, которая в наше время успешно применена к позвоночным животным; П.Г.Светлова (1892–1974), высказавшего глубокие идеи о взаимоотношении целостных и«элементаристических» подходов в биологии развития; Г.А.Шмидта, исследовавшего ряд проблем сравнительной эмбриологии беспозвоночных и позвоночных животных; Б.П.Токина, развивающего учение о соматических эмбриогенезах, а также Г.А.Кнорре, Л.Я.Бляхера и Г.В.Лопашова. В отношении формирования современных представлений об индивидуальном развитии большую роль сыграли цитологические и генетические исследования, которые к 30–40-м годам все более тесно смыкались с экспериментально-эмбриологическим направлением.
Бесполое размножение
Бесполое размножение – процесс возникновения дочерних особей из одной или группы соматических клеток материнского организма. Этот способ размножения более древний. В его основе лежит митотическое деление клеток. Значение бесполого размножения заключается в быстром увеличении числа особей, почти не различающихся между собой. Различают следующие формы бесполого размножения. Деление надвое – приводит к возникновению из одного родительского организма двух дочерних. Является преобладающей формой деления у прокариот и простейших. Различные одноклеточные животные делятся по-разному. Так, жгутиковые делятся продольно, а инфузории – поперечно. Такое деление встречается и у многоклеточных животных – кишечнополостных (продольное деление у медуз) и червей (поперечное деление у кольчатых червей). Множественное деление (шизогония) – встречается среди простейших, в том числе у паразитов человека (малярийный плазмодий). Почкование – на теле материнского организма возникает скопление клеток, которое растет и постепенно приобретает сходство с материнской особью. Затем дочерняя особь отделяется и начинает вести самостоятельное существование. Такое размножение распространено среди низших многоклеточных (губки, кишечнополостные, мшанки, некоторые черви и оболочники). Иногда дочерние особи не отделяются полностью от родительской, что приводит к образованию колоний. Фрагментация – происходит распад тела многоклеточного организма на части, которые в дальнейшем превращаются в самостоятельные особи (плоские черви, иглокожие). Спорами – дочерний организм развивается из специализированной клетки-споры. Бесполое размножение наблюдается у животных с относительно низким уровнем морфофизиологической организации. А у растений распространенность бесполого размножения настолько велика, что может считаться их характерной особенностью. Как низшие (водоросли), так и высшие (споровые, голосеменные, покрытосеменные) растения размножаются бесполым путем. Различают две основные формы бесполого размножения растений: вегетативное размножение и спорообразование. Вегетативное размножение одноклеточных растений осуществляется простым делением одной клетки на две. У грибов формы его более разнообразны – спорообразование (плесневые грибы, шляпочные) и почкование (дрожжи). У покрытосеменных растений вегетативное размножение происходит за счет вегетативных (неполовых) органов – корня, стебля, листа. При вегетативном размножении используются видоизменения этих органов, такие как клубни (картофель, георгин), корневища (флокс), плети (земляника), луковицы (лук, тюльпан), корневые отпрыски (малина, вишня). Многие растения размножаются черенками и отводками. Спорообразование – собственно бесполое размножение растений. Оно осуществляется специализированными клетками – спорами, которые образуются чаще всего в органах бесполого размножения – спорангиях. Для низших растений типично размножение зооспорами, которые образуются митотически, являются диплоидными и способны воспроизводить новую особь, сходную с материнской. Споры высших растений обычно гаплоидны, так как образуются в результате мейотического деления. Эти споры не могут воспроизвести материнскую особь. Они служат для формирования такой жизненной стадии растения, как гаметофит, который образует половые клетки – гаметы. У растений и низкоорганизованных животных, которым характерно бесполое размножение, обычно происходит чередование бесполой и половой форм размножения в их жизненном цикле. Различные формы бесполого размножения представлены в таблице 2. Половое размножение
Половое размножение – наблюдается в жизненных циклах всех основных групп организмов. Распространенность полового размножения объясняется тем, что оно обеспечивает значительное генетическое разнообразие и, следовательно, фенотипическую изменчивость потомства. В основе полового размножения лежит половой процесс, суть которого сводится к объединению в наследственном материале для развития потомка генетической информации от двух разных источников – родителей. Одной из форм полового процесса является конъюгация. При этом происходит временное соединение двух особей с целью обмена (рекомбинации) наследственным материалом, например, у инфузорий. В результате появляются особи генетически отличные от родительских организмов, которые в дальнейшем осуществляют бесполое размножение. Число инфузорий после конъюгации не изменяется, поэтому говорить в прямом смысле о размножении в этом случае нельзя. У простейших половой процесс может осуществляться и в форме копуляции – слияния двух особей в одну, объединение и рекомбинация наследственного материала. Далее такая особь размножается делением. На определенном этапе эволюции у многоклеточных организмов половой процесс как способ обмена генетической информацией между особями в пределах вида оказался связанным с размножением. При половом размножении возникающие новые особи обычно отличаются от родительских и друг от друга комбинацией аллелей генов. Новые сочетания хромосом и генов проявляются у потомков новым сочетанием признаков. В результате возникает большое разнообразие особей в пределах одного вида. Таким образом, биологическое значение полового размножения заключается не только в самовоспроизведении, но и в обеспечении исторического развития видов, то есть жизни как таковой. Это позволяет считать половое размножение биологически более прогрессивным, чем бесполое. Для участия в половом размножении в родительских организмах вырабатываются гаметы – клетки, специализированные к обеспечению генеративной функции. Слияние материнской и отцовской гамет приводит к возникновению зиготы – клетки, представляющей собой дочернюю особь на первой, наиболее ранней стадии индивидуального развития. У некоторых организмов зигота образуется в результате объединения гамет, которые не отличаются по строению – явление изогамии. У большинства же видов половые клетки по структурным и функциональным признакам делятся на материнские (яйцеклетки) и отцовские (сперматозоиды).
Таблица 2 Формы бесполого размножения
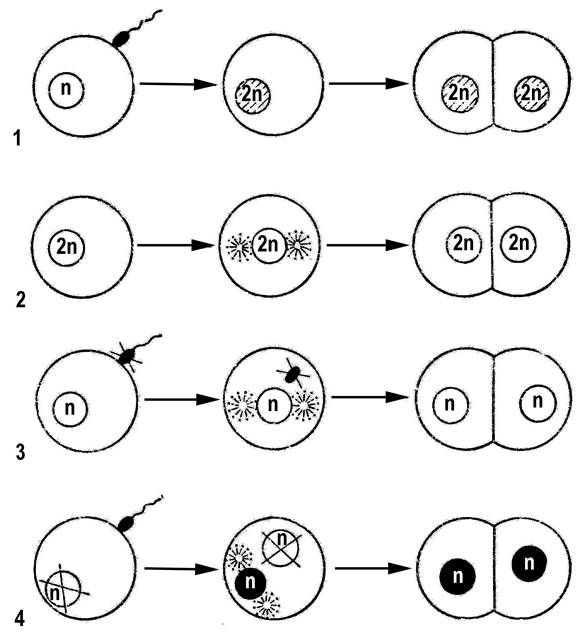
Яйцеклетки и сперматозоиды вырабатываются разными организмами – женскими (самками) и мужскими (самцами). В разделении гамет на яйцеклетки и сперматозоиды и особей на самок и самцов заключается явление половогодиморфизма. Наличие этого явления в природе отражает различия в задачах, решаемых в процессе полового размножения мужской или женской гаметой, самцом или самкой. Иногда развитие дочернего организма происходит из неоплодотворенной яйцеклетки. Это явление называют девственнымразвитием или партеногенезом. Источником наследственного материала для развития потомка в этом случае обычно служит ДНК яйцеклетки – явление гиногенеза. Реже наблюдается андрогенез – развитие потомка из клетки с цитоплазмой ооцита и ядром сперматозоида. Ядро женской гаметы в случае андрогенеза погибает. Обязательный партеногенез является измененной формой полового размножения в эволюции некоторых видов животных. Например, у пчел он используется как механизм генотипического определения пола: женские особи (рабочие пчелы и царицы) развиваются из оплодотворенных яйцеклеток, а мужские (трутни) – партеногенетически. Партеногенез включен в жизненный цикл многих паразитических видов. Он обеспечивает рост численности особей в условиях, при которых затрудняется встреча партнеров противоположного пола. При партеногенезе, как и при типичном половом размножении, развиваются особи с диплоидными соматическими клетками. Восстановление диплоидного набора хромосом происходит обычно путем слияния ооцита и редукционного тельца во втором делении мейоза. Помимо естественного партеногенеза существует искусственный, который вызывается воздействием различных неспецифических факторов – нагревание, облучение, механическое воздействие иглой, действие кислот, осуществляемых человеком. Искусственный партеногенез позволяет получать поколения, представленные организмами только одного пола. Это имеет важнейшее практическое значение в сельском хозяйстве. Ярким свидетельством этого являются работы Б.Л.Астаурова по искусственному партеногенезу тутового шелкопряда. Ядра яйцеклеток инактивируют прогревом и облучением яиц. Затем яйца оплодотворяют. Из ядер двух проникнувших в яйцо сперматозоидов восстанавливается диплоидный набор хромосом, при этом из таких яиц развиваются только самцы. Этот результат имеет простое объяснение с позиций цитогенетики. Самцы тутового шелкопряда содержат в кариотипе две ХХ-хромосомы, поэтому все сперматозоиды в своем гаплоидном наборе несут только Х-хромосому. Следовательно, при слиянии ядер двух сперматозоидов обязательно образуется хромосомный набор самца ХХ. Коконы самцов содержат больше шелка, чем коконы самок. В результате удается получать экономическую выгоду. У некоторых видов наблюдается полиэмбриония – бесполое размножение зародыша, образовавшегося путем полового размножения. Например, у броненосцев на стадии бластулы происходит разделение клеточного материала первоначально одного зародыша между 4–8 зародышами, из которых в последствии развиваются полноценные особи. В результате полиэмбрионии у человека рождаются однояйцовые близнецы. Различные типы полового размножения животных и растений представлены на рис. 2.
Рис. 2. Типы полового размножения: 1 – нормальное оплодотворение; 2 – партеногенез; 3 – гиногенез; 4 – андрогенез ЛЕКЦИЯ 3
ПРОГЕНЕЗ ПЛАН
1. Сперматогенез. 2. Овогенез. 3. Отличия половых клеток от соматических.
Сперматогенез Началу индивидуального развития предшествует возникновение половых клеток, т.е. гаметогенез, который можно рассматривать прогенезом при индивидуальном развитии. Процесс образования мужских половых клеток – сперматогенез. В результате образуются сперматозоиды. Соматические клетки, достигнув определенного зрелого физиологического состояния, делятся митотически (иногда путем амитоза), половые же клетки в своем развитии проходят особые фазы превращения, пока не созреют и не станут способными к оплодотворению. Разница эта имеет глубокий биологический смысл. Соматические клетки должны сохранить всю сумму наследственной информации в ходе делений, чтобы дочерние клетки оставались такими же, как и материнские. Передача информации обеспечивается в ходе митоза точным распределением хромосом между делящимися клетками: число хромосом, их биологическая структура, содержание ДНК и, следовательно, заключенная в ней наследственная информация сохраняются в ряду клеточных поколений, обеспечивая постоянство строения особи и вида. При оплодотворении ядра мужской и женской половых клеток объединяются в общее ядро, и если бы хромосом в каждой было столько же, сколько в соматических клетках, то в зиготе оно удваивалось бы, и такое удвоенное количество
|
||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-06-22; просмотров: 1313; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.108 (0.054 с.) |

 ISBN 5-87978-288-3
ISBN 5-87978-288-3