Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Становление биологии индивидуального развитияСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Еще в 20-е годы Н.К.Кольцов создал целое направление теоретических и экспериментальных исследований физико-химической природы и механизмов функционирования хромосомы как одного из клеточных органоидов. Его можно считать основоположником биологии клетки, ему принадлежит заслуга в создании экспериментальной и физико-химической биологии в СССР. Можно только удивляться, насколько верными, далеко опережающими свое время были методологические принципы, которыми руководствовался Н.К.Кольцов, призывавший к аналитическому исследованию клетки вплоть до молекул, но с обязательным синтезом полученных данных для правильного понимания роли элементарных структур в жизнедеятельности и в развитии целостной системы (клетки). Комплексный подход, глубокое чутье и прозорливость позволили ему в конце 30-х – начале 40-х годов высказать идеи об устройстве и репродукции хромосомы как самовоспроизводящейся гигантской белковой молекулы, способной строить себе подобную из элементов окружающей среды. Эти идеи предвосхитили будущие открытия принципов матричного синтеза и механизмов редупликации. Он придавал большое значение конкретным физико-химическим условиям, в которых функционирует хромосома,– цитоплазме, призывая использовать достижения цитологии, биохимии и генетики для изучения обмена веществ между ядром и цитоплазмой. В соответствии с современными представлениями Кольцов считал, что ядерный материал может выходить в цитоплазму, придавая ей наследственные потенции («материнский эффект»). Широта биологического подхода, понимание важности объединения достижений цитологии, биохимии, генетики, экспериментальной эмбриологии и физико-химических направлений, необходимости сочетания исследований на молекулярном уровне и на уровне целостной системы для разработки фундаментальных проблем биологии клетки – все это позволяет считать Н.К.Кольцова одним из создателей современной биологии индивидуального развития. Идеи и направления работ Н.К.Кольцова успешно развивал его ученик – акад. Б.Л.Астауров, который уже в 30-е годы говорил о необходимости изучения ядерно-цитоплазматических отношений как основы деятельности генетического аппарата. В числе первых Б.Л.Астауров пришел к выводу о том, что проявление гена – это сложный и отнюдь не заданный в полной мере процесс. Заметим, что только те биологи, которые своевременно оценили соотношение между закономерным и случайным в процессе развития и, в частности, в процессе реализации генетической информации, были ближе всех к современной диалектико-материалистической методологии и современным научным представлениям. Б.Л.Астауров был одним из них. Заслуга Б.Л.Астаурова состоит в том, что он первый разработал точные экспериментально-генетические подходы к изучению роли ядра и цитоплазмы в развитии признаков животных. Методы искусственного получения партеногенетического, андрогенетического и гиногенетического потомства, которые он начал разрабатывать еще в 40-е годы, вошли в золотой фонд научно-прикладных достижений современной биологии. В 50–60-е годы началось целенаправленное и широкое экспериментальное исследование роли ядерно-цитоплазматических отношений в развитии. Усовершенствование микрохирургической техники позволило существенно продвинуть разработку этого вопроса и использовать методы пересадок ядер дифференцированных клеток в яйцеклетки с целью выяснения их потенций в новом окружении. Наиболее важных результатов здесь достигли американские исследователи Т.Кинг, Р.Бриггс и английский эмбриолог Дж.Гердон. В те же годы продолжалось исследование индукционных связей (С.Тойвонен, Л.Саксен, П.Ньюкуп, К.Гробстайн и др.), был открыт целый ряд так называемых вторичных индукций, интенсивно исследовалась их природа. Логика работ в этом направлении постепенно вела исследователей от межорганных к межклеточным взаимодействиям. Изучение механизмов межклеточных взаимодействий и их роли в морфогенезе, дифференцировке и в осуществлении индукционных связей – одна из наиболее важных отраслей современной биологии, в которой экспериментальная эмбриология взаимодействует с цитологией, и молекулярной биологией. В 60-х годах произошел окончательный синтез экспериментально-эмбриологических, экспериментально-цитологических, генетических и физико-химических направлений, связанных с изучением процессов развития, и возникла самостоятельная область – биология индивидуального развития. ЛЕКЦИЯ 2
РАЗМНОЖЕНИЕ ОРГАНИЗМОВ ПЛАН
1. Размножение – свойство живых организмов. 2. Бесполое размножение. 3. Половое размножение.
Размножение – свойство живых организмов
Среди многообразных проявлений жизнедеятельности организмов особая роль принадлежит размножению. В определенном смысле существование самого организма является подготовкой к выполнению им главной биологической задачи – участию в размножении. В основе способности организмов к размножению лежат определенные клеточные механизмы – митоз, мейоз и оплодотворение. Эти процессы обеспечивают сохранение и передачу основных признаков данного вида организмов от родительского поколения к потомкам. Способность к размножению является неотъемлемым свойством живых существ. Благодаря размножению осуществляется материальная преемственность и непрерывность поколений, а следовательно, непрерывность и преемственность жизни. Таким образом, биологическое значение размножения состоит в том, что оно обеспечивает сменупоколений определенного вида организмов. При размножении в ряду поколений осуществляется передача генетического материала (ДНК), т.е. определенной, специфической для данного вида биологической информации. Наследственные изменения организмов в ряду поколений приводят к изменчивости вида и к эволюции. Размножение возникло на самом раннем этапе исторического развития органического мира одновременно с образованием клетки. В зависимости от характера клеточного материала, используемого в целях размножения, выделяют различные способы и формы размножения. В живой природе встречаются два главных способа размножения: бесполое и половое. Общая характеристика бесполого и полового размножения представлена в таблице 1.
Таблица 1 Общая характеристика бесполого и полового размножения
Бесполое размножение
Бесполое размножение – процесс возникновения дочерних особей из одной или группы соматических клеток материнского организма. Этот способ размножения более древний. В его основе лежит митотическое деление клеток. Значение бесполого размножения заключается в быстром увеличении числа особей, почти не различающихся между собой. Различают следующие формы бесполого размножения. Деление надвое – приводит к возникновению из одного родительского организма двух дочерних. Является преобладающей формой деления у прокариот и простейших. Различные одноклеточные животные делятся по-разному. Так, жгутиковые делятся продольно, а инфузории – поперечно. Такое деление встречается и у многоклеточных животных – кишечнополостных (продольное деление у медуз) и червей (поперечное деление у кольчатых червей). Множественное деление (шизогония) – встречается среди простейших, в том числе у паразитов человека (малярийный плазмодий). Почкование – на теле материнского организма возникает скопление клеток, которое растет и постепенно приобретает сходство с материнской особью. Затем дочерняя особь отделяется и начинает вести самостоятельное существование. Такое размножение распространено среди низших многоклеточных (губки, кишечнополостные, мшанки, некоторые черви и оболочники). Иногда дочерние особи не отделяются полностью от родительской, что приводит к образованию колоний. Фрагментация – происходит распад тела многоклеточного организма на части, которые в дальнейшем превращаются в самостоятельные особи (плоские черви, иглокожие). Спорами – дочерний организм развивается из специализированной клетки-споры. Бесполое размножение наблюдается у животных с относительно низким уровнем морфофизиологической организации. А у растений распространенность бесполого размножения настолько велика, что может считаться их характерной особенностью. Как низшие (водоросли), так и высшие (споровые, голосеменные, покрытосеменные) растения размножаются бесполым путем. Различают две основные формы бесполого размножения растений: вегетативное размножение и спорообразование. Вегетативное размножение одноклеточных растений осуществляется простым делением одной клетки на две. У грибов формы его более разнообразны – спорообразование (плесневые грибы, шляпочные) и почкование (дрожжи). У покрытосеменных растений вегетативное размножение происходит за счет вегетативных (неполовых) органов – корня, стебля, листа. При вегетативном размножении используются видоизменения этих органов, такие как клубни (картофель, георгин), корневища (флокс), плети (земляника), луковицы (лук, тюльпан), корневые отпрыски (малина, вишня). Многие растения размножаются черенками и отводками. Спорообразование – собственно бесполое размножение растений. Оно осуществляется специализированными клетками – спорами, которые образуются чаще всего в органах бесполого размножения – спорангиях. Для низших растений типично размножение зооспорами, которые образуются митотически, являются диплоидными и способны воспроизводить новую особь, сходную с материнской. Споры высших растений обычно гаплоидны, так как образуются в результате мейотического деления. Эти споры не могут воспроизвести материнскую особь. Они служат для формирования такой жизненной стадии растения, как гаметофит, который образует половые клетки – гаметы. У растений и низкоорганизованных животных, которым характерно бесполое размножение, обычно происходит чередование бесполой и половой форм размножения в их жизненном цикле. Различные формы бесполого размножения представлены в таблице 2. Половое размножение
Половое размножение – наблюдается в жизненных циклах всех основных групп организмов. Распространенность полового размножения объясняется тем, что оно обеспечивает значительное генетическое разнообразие и, следовательно, фенотипическую изменчивость потомства. В основе полового размножения лежит половой процесс, суть которого сводится к объединению в наследственном материале для развития потомка генетической информации от двух разных источников – родителей. Одной из форм полового процесса является конъюгация. При этом происходит временное соединение двух особей с целью обмена (рекомбинации) наследственным материалом, например, у инфузорий. В результате появляются особи генетически отличные от родительских организмов, которые в дальнейшем осуществляют бесполое размножение. Число инфузорий после конъюгации не изменяется, поэтому говорить в прямом смысле о размножении в этом случае нельзя. У простейших половой процесс может осуществляться и в форме копуляции – слияния двух особей в одну, объединение и рекомбинация наследственного материала. Далее такая особь размножается делением. На определенном этапе эволюции у многоклеточных организмов половой процесс как способ обмена генетической информацией между особями в пределах вида оказался связанным с размножением. При половом размножении возникающие новые особи обычно отличаются от родительских и друг от друга комбинацией аллелей генов. Новые сочетания хромосом и генов проявляются у потомков новым сочетанием признаков. В результате возникает большое разнообразие особей в пределах одного вида. Таким образом, биологическое значение полового размножения заключается не только в самовоспроизведении, но и в обеспечении исторического развития видов, то есть жизни как таковой. Это позволяет считать половое размножение биологически более прогрессивным, чем бесполое. Для участия в половом размножении в родительских организмах вырабатываются гаметы – клетки, специализированные к обеспечению генеративной функции. Слияние материнской и отцовской гамет приводит к возникновению зиготы – клетки, представляющей собой дочернюю особь на первой, наиболее ранней стадии индивидуального развития. У некоторых организмов зигота образуется в результате объединения гамет, которые не отличаются по строению – явление изогамии. У большинства же видов половые клетки по структурным и функциональным признакам делятся на материнские (яйцеклетки) и отцовские (сперматозоиды).
Таблица 2 Формы бесполого размножения
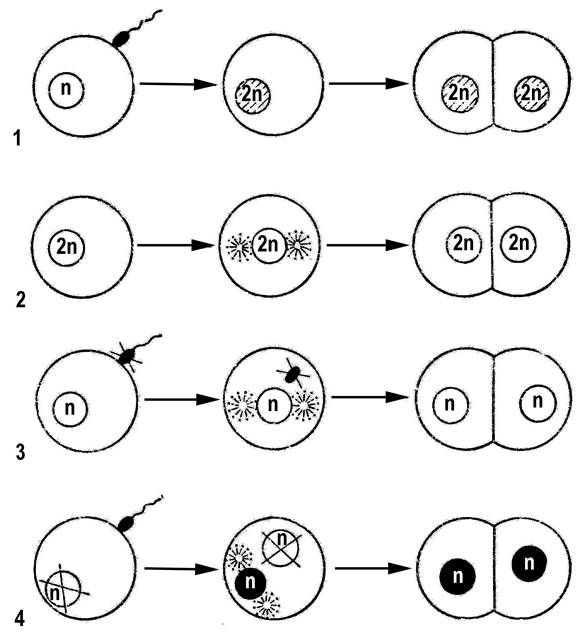
Яйцеклетки и сперматозоиды вырабатываются разными организмами – женскими (самками) и мужскими (самцами). В разделении гамет на яйцеклетки и сперматозоиды и особей на самок и самцов заключается явление половогодиморфизма. Наличие этого явления в природе отражает различия в задачах, решаемых в процессе полового размножения мужской или женской гаметой, самцом или самкой. Иногда развитие дочернего организма происходит из неоплодотворенной яйцеклетки. Это явление называют девственнымразвитием или партеногенезом. Источником наследственного материала для развития потомка в этом случае обычно служит ДНК яйцеклетки – явление гиногенеза. Реже наблюдается андрогенез – развитие потомка из клетки с цитоплазмой ооцита и ядром сперматозоида. Ядро женской гаметы в случае андрогенеза погибает. Обязательный партеногенез является измененной формой полового размножения в эволюции некоторых видов животных. Например, у пчел он используется как механизм генотипического определения пола: женские особи (рабочие пчелы и царицы) развиваются из оплодотворенных яйцеклеток, а мужские (трутни) – партеногенетически. Партеногенез включен в жизненный цикл многих паразитических видов. Он обеспечивает рост численности особей в условиях, при которых затрудняется встреча партнеров противоположного пола. При партеногенезе, как и при типичном половом размножении, развиваются особи с диплоидными соматическими клетками. Восстановление диплоидного набора хромосом происходит обычно путем слияния ооцита и редукционного тельца во втором делении мейоза. Помимо естественного партеногенеза существует искусственный, который вызывается воздействием различных неспецифических факторов – нагревание, облучение, механическое воздействие иглой, действие кислот, осуществляемых человеком. Искусственный партеногенез позволяет получать поколения, представленные организмами только одного пола. Это имеет важнейшее практическое значение в сельском хозяйстве. Ярким свидетельством этого являются работы Б.Л.Астаурова по искусственному партеногенезу тутового шелкопряда. Ядра яйцеклеток инактивируют прогревом и облучением яиц. Затем яйца оплодотворяют. Из ядер двух проникнувших в яйцо сперматозоидов восстанавливается диплоидный набор хромосом, при этом из таких яиц развиваются только самцы. Этот результат имеет простое объяснение с позиций цитогенетики. Самцы тутового шелкопряда содержат в кариотипе две ХХ-хромосомы, поэтому все сперматозоиды в своем гаплоидном наборе несут только Х-хромосому. Следовательно, при слиянии ядер двух сперматозоидов обязательно образуется хромосомный набор самца ХХ. Коконы самцов содержат больше шелка, чем коконы самок. В результате удается получать экономическую выгоду. У некоторых видов наблюдается полиэмбриония – бесполое размножение зародыша, образовавшегося путем полового размножения. Например, у броненосцев на стадии бластулы происходит разделение клеточного материала первоначально одного зародыша между 4–8 зародышами, из которых в последствии развиваются полноценные особи. В результате полиэмбрионии у человека рождаются однояйцовые близнецы. Различные типы полового размножения животных и растений представлены на рис. 2.
Рис. 2. Типы полового размножения: 1 – нормальное оплодотворение; 2 – партеногенез; 3 – гиногенез; 4 – андрогенез ЛЕКЦИЯ 3
ПРОГЕНЕЗ ПЛАН
1. Сперматогенез. 2. Овогенез. 3. Отличия половых клеток от соматических.
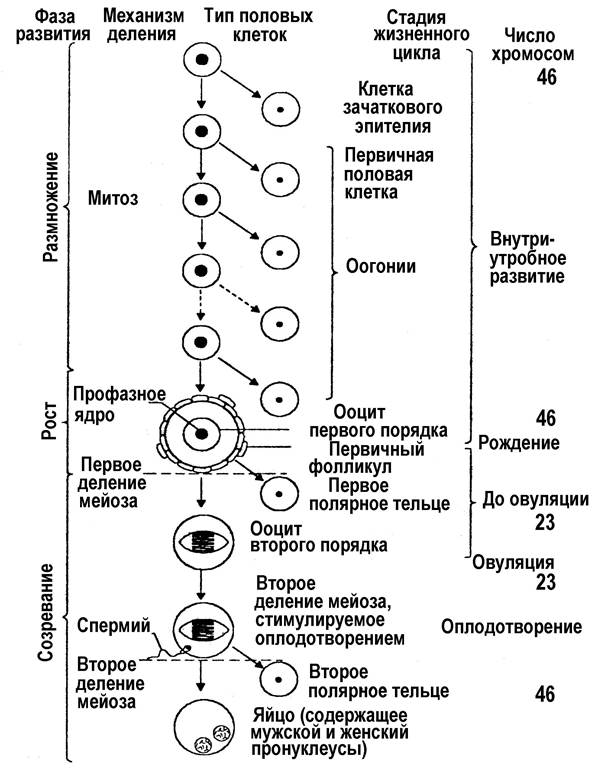
Сперматогенез Началу индивидуального развития предшествует возникновение половых клеток, т.е. гаметогенез, который можно рассматривать прогенезом при индивидуальном развитии. Процесс образования мужских половых клеток – сперматогенез. В результате образуются сперматозоиды. Соматические клетки, достигнув определенного зрелого физиологического состояния, делятся митотически (иногда путем амитоза), половые же клетки в своем развитии проходят особые фазы превращения, пока не созреют и не станут способными к оплодотворению. Разница эта имеет глубокий биологический смысл. Соматические клетки должны сохранить всю сумму наследственной информации в ходе делений, чтобы дочерние клетки оставались такими же, как и материнские. Передача информации обеспечивается в ходе митоза точным распределением хромосом между делящимися клетками: число хромосом, их биологическая структура, содержание ДНК и, следовательно, заключенная в ней наследственная информация сохраняются в ряду клеточных поколений, обеспечивая постоянство строения особи и вида. При оплодотворении ядра мужской и женской половых клеток объединяются в общее ядро, и если бы хромосом в каждой было столько же, сколько в соматических клетках, то в зиготе оно удваивалось бы, и такое удвоенное количество переходило бы во все клетки развивающегося зародыша. В дальнейшем, при развитии половых клеток следующих поколений молодых организмов будет происходить последовательное накопление хромосом в клетках, и вид не мог бы сохранить неизменными свои наследственные особенности. Кроме того, постепенно нарушался бы ядерно-плазменный коэффициент в пользу ядра, и через несколько поколений наступил бы такой момент, когда добавление хромосом в ядро вело бы к неизбежной гибели клетки. В результате, оплодотворение стало бы служить не для сохранения, а для уничтожения организмов. Однако этого не случается, так как в процесс гаметогенеза включены два особых деления, в ходе которых число хромосом в ядрах как мужской, так и женской половой клетки уменьшается вдвое. Внутриклеточные процессы, связанные с уменьшением числа хромосом, составляют существо созревания половых клеток – существо мейоза. При оплодотворении половинное количество хромосом ядер клеток отца и половинное количество хромосом ядер клеток матери объединяются, и в зиготе восстанавливается свойственный данному виду набор хромосом. В сперматогенезе различают 4 периода: размножение, рост, созревание (мейоз) и формирование (рис. 3). В период размножения исходные недифференцированные половые клетки – сперматогонии, или гонии делятся путем обычного митоза. Проделав несколько таких делений, они вступают в период роста. На этой стадии их называют сперматоцитами I порядка (или цитами I). Они усиленно ассимилируют питательные вещества, укрупняются, претерпевают глубокую физико-химическую перестройку, в результате которой подготавливаются к третьему периоду – созреванию, или мейозу. В мейозе сперматоциты I проходят два процесса клеточного деления. В первом делении (редукционном) происходит уменьшение числа хромосом (редукция). В результате из одного цита I возникает две равновеликие клетки – сперматоциты II порядка, или циты II. Затем наступает второе деление созревания. Оно протекает как обычный соматический митоз, но при гаплоидном числе хромосом. Такое деление называется эквационным («эквацио» – равенство), так как образуются две тождественные, т.е. полностью равноценные клетки, которые называются сперматидами. В четвертом периоде – формирования – округлая сперматида приобретает форму зрелой мужской половой клетки: у нее вырастает жгутик, уплотняется ядро, образуется оболочка. В результате всего процесса сперматогенеза из каждой исходной недифференцированной сперматогонии получается 4 зрелых половых клетки, содержащих по гаплоидному набору хромосом. На рис. 4 представлена схема процессов сперматогенеза и спермиогенеза у человека. Сперматогенез происходит в извитых семенных канальцах семенников. Развитие сперматозоидов начинается в период пренатального развития при закладке генеративных тканей, затем возобновляется в период наступления половозрелости и продолжается до старости. В период размножения происходит ряд следующих друг за другом митозов, в результате которых происходит увеличение количества клеток, называемых сперматогониями. Некоторые сперматогонии вступают в период роста и называются сперматоцитамиI порядка. Период роста соответствует периоду интерфазы клеточного цикла, в которой происходит удвоение наследственного материала сперматоцитов I порядка (2n4С), и затем они вступают в профазу I мейотического деления. Во время профазы I происходит конъюгация гомологичных хромосом и обмен между гомологичными хроматидами (кроссинговер). Кроссинговер имеет важное генетическое значение, так как приводит к возникновению генетических различий между индивидуумами.
Рис. 3. Схема гаметогенеза: 1-й – период размножения: клетки делятся митотически, набор хромосом в них 2n; 2-й – период роста: накопление в клетках питательных веществ, набор хромосом них 2n; 3-й – период созревания – мейоз: а)1-е, или редукционное, деление, образование из диплоидных клеток с набором хромосом, равным 2n, клеток с гаплоидным набором, равным n; б) 2-е деление мейоза, протекает как митоз, но в клетках с гаплоидным набором хромосом; 4-й – период формирования – имеет место только в сперматогенезе
Период созревания протекает в два этапа, что соответствует I мейотическому (редукционному) и II мейотическому (эквационному) делениям. При этом из одного сперматоцита I порядка сначала получаются два сперматоцита II порядка (1n2С), затем 4 сперматиды (1n1С). Сперматиды отличаются друг от друга набором хромосом: они все содержат по 22 аутосомы, но половина клеток содержит Х-хромосому, а другая половина – Y-хромосому. Аутосомы отличаются между собой и от родительских различным сочетанием аллелей, поскольку произошел обмен во время кроссинговера. В период формирования количество клеток и число хромосом в них не меняется, т.к. в этот период из 4 сперматид формируется 4 сперматозоида, в которых происходит морфологическая реорганизация клеточных структур, формируется хвост. У человека эта фаза продолжается 14 дней. Мужские половые клетки не развиваются одиночно, они растут в клонах и объединены между собой цитоплазматическими мостиками. Цитоплазматические мостики имеются между сперматогониями, сперматоцитами и сперматидами. В конце фазы формирования сперматозоиды освобождаются от цитоплазматических мостиков. У человека максимум дневной продуктивности сперматозоидов 108, продолжительность существования сперматозоида во влагалище до 2,5 ч, а в шейке матки до 48 ч.
Рис. 4. Схема процессов сперматогенеза и спермиогенеза у человека (по Bloom and Fawcett, 1975) Овогенез
Процесс развития женских половых клеток называется овогенезом (оогенезом). В отличие от сперматогенеза он имеет некоторые особенности. Ход овогенеза и его отличия от развития мужских гамет показаны на рис. 3. В овогенезе различают 3 периода: размножение, рост и созревание. Недифференцированные женские половые клетки – овогонии – размножаются так же, как и сперматогонии, путем обычного митоза. После деления они становятся овоцитами I порядка и переходят в период роста. Рост овоцитов длится очень долго – недели, месяцы и даже годы. В периоде роста различают два этапа: малый, или медленный рост, когда ассимилируются новые вещества и ими обогащается преимущественно цитоплазма, и большой, или быстрый рост, когда в клетке накапливается питательный желток. Глубокие изменения в периоде роста претерпевает и ядро, оно сильно набухает, содержимое его как бы расплывается. Размеры клеток колоссально возрастают (например, икринки окуня увеличиваются почти в миллион раз). Затем овоцит I порядка вступает в период созревания, или мейоз. Здесь тоже совершаются редукционное и эквационное деления. Процессы деления в ядре протекают так же, как при мейозе сперматоцитов, но судьба цитоплазмы совершенно иная. При редукционном делении одно ядро увлекает с собой бóльшую часть цитоплазмы, а на долю другого остается лишь незначительная ее часть. Поэтому образуется только одна полноценная клетка – овоцит II порядка, и вторая крошечная – направительное, или редукционное, тельце, которое может делиться на два редукционных тельца. При втором, эквационном делении несимметричное распределение цитоплазмы повторяется и опять образуется одна крупная клетка – овотида и третье полярное тельце. Овотида по составу ядра и функционально является вполне зрелой половой клеткой. Период формирования, в отличие от сперматогенеза, в овогенезе отсутствует. Таким образом, в овогенезе из одной овогонии возникает только одна зрелая яйцеклетка. Полярные тельца остаются недоразвитыми и вскоре погибают и фагоцитируются другими клетками. Зрелые женские гаметы называют яйцеклетками или яйцами, а отложенные в воду – икрой. Особенности оогенеза у человека представлены на рис. 5. Развитие женских половых клеток происходит в яичниках. Период размножения наступает у оогоний еще у зародыша и прекращается к моменту рождения девочки.
Рис. 5. Схема процесса оогенеза у человека (по Н.Грин, 1990) Период роста при оогенезе более продолжительный, т.к. кроме подготовки к мейозу осуществляется накопление запаса питательных веществ, которые будут необходимы в дальнейшем для первых дроблений зиготы. В фазе малого роста происходит образование большого количества разных типов РНК. Быстрое накопление РНК происходит за счет специального механизма – амплификациигенов (множественное копирование отдельных участков ДНК, кодирующих рибосомную РНК). Быстрое увеличение мРНК идет за счет образования хромосом типа «ламповых щеток». В результате образуется более тысячи дополнительных ядрышек, которые являются необходимой структурой для синтеза рРНК, из которой впоследствии формируются рибосомы, участвующие в синтезе белка. В этот же период в ооците происходят мейотические преобразования хромосом, характерные для осуществления профазы первого деления. В период большого роста фолликулярные клетки яичника образуют несколько слоев вокруг ооцита I порядка, что способствует переносу питательных веществ, синтезированных в других местах, в цитоплазму ооцита. У человека период роста ооцитов может составлять 12–50 лет. После завершения периода роста ооцит I порядка вступает в период созревания. В периоде созревания ооцитов (также как и при сперматогенезе) осуществляется мейотическое деление клеток. При первом редукционном делении из ооцита I порядка образуется один ооцит II порядка (1n2С) и одно полярное тельце (1n2С). При втором эквационном делении из ооцита II порядка образуется созревшая яйцеклетка (1n1С), сохранившая почти все накопленные вещества в цитоплазме, и второе полярное тельце маленьких размеров (1n1С). В это же время происходит деление первого полярного тельца, дающего начало двум вторым полярным тельцам (1n1С). В результате при оогенезе получается 4 клетки, из которых только одна станет в дальнейшем яйцеклеткой, а остальные 3 (полярные тельца) редуцируются. Биологическая значимость этого этапа оогенеза – сохранить все накопленные вещества цитоплазмы около одного гаплоидного ядра для обеспечения нормального питания и развития оплодотворенной яйцеклетки. При оогенезе у женщин на стадии второй метафазы образуется блок, который снимается во время оплодотворения, и фаза созревания заканчивается только после проникновения сперматозоида в яйцеклетку. Процесс оогенеза у женщин – это циклический процесс, повторяющийся примерно через каждые 28 дней (начиная с периода роста и заканчивая период только после оплодотворения). Этот цикл называется менструальным. Отличительные особенности сперматогенеза и овогенеза у человека представлены в таблице 3.
Таблица 3
|
|||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-06-22; просмотров: 1250; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.20 (0.018 с.) |