Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
НОВОСИБИРСКий государственныйСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
НОВОСИБИРСКий государственный МЕДИЦИНСКий университет КАФЕДРА НОРМАЛЬНОЙ ФИЗИОЛОГИИ
УЧЕБНО-МЕТОДИЧЕСКОЕ ПОСОБИЕ
Общие закономерности гормональной регуляции
НОВОСИБИРСК 2007 г.
НОВОСИБИРСКий государственный МЕДИЦИНСКий университет КАФЕДРА НОРМАЛЬНОЙ ФИЗИОЛОГИИ
УЧЕБНО-МЕТОДИЧЕСКОЕ ПОСОБИЕ ПО КУРСУ НОРМАЛЬНОЙ ФИЗИОЛОГИИ
«Общие закономерности гормональной регуляции»
Автор-составитель: профессор кафедры нормальной физиологии НГМУ, д. б. н. Н.Б. Пиковская
НОВОСИБИРСК 2007 г.
Методическое пособие по курсу нормальной физиологии к разделу: Физиология эндокринной системы «Общие закономерности гормональной регуляции» Новосибирская Государственная медицинская академия, 2007. - 76 с. Методическое пособие предназначено для использования в курсе практических занятий по нормальной физиологии студентами второго курса всех факультетов. Утверждено Центральной комиссией методического совета НГМА - протокол № от 2007 г.
Автор-составитель: профессор кафедры нормальной физиологии НГМА, д. б. н. Н.Б. Пиковская
Рецензенты: Зав. кафедрой педагогики и медицинской психологии НГМУ, доцент Г. В. Безродная Профессор кафедры биохимии НГМУ, д.м.н., профессор Н.Н. Маянская
ã Новосибирский государственный медицинский университет
Предисловие Это учебно-методическое пособие составлено в соответствии с лекционным курсом, который в течение последних лет читается на кафедре нормальной физиологии. Прекрасно понимая невозможность в лекционном курсе или коротком пособии изложить все физиологические эффекты гормонов, мы сосредоточили свое внимание на самых общих закономерностях гормональной регуляции, в надежде на то, что эти базовые знания позволят будущим медикам углубить их при необходимости. В своем курсе мы оказались от традиционного изложения свойств отдельных гормонов в соответствии с анатомической принадлежностью, отдав предпочтение общебиологическому значению гормональной регуляции. Поэтому, остановившись коротко на общих закономерностях регуляции секреции гормонов, более подробно рассматриваем значение гормонов в процессах роста и развития организма, регуляции основных параметров гомеостаза (глюкоза, кальций, натрий). Особое внимание и в лекционном курсе, и в пособии уделяется роли гормонов в адаптации, стрессу, как основному механизму адаптации и повреждающим эффектам высоких доз гормонов при длительном стрессе. Изложение физиологии эндокринной регуляции в таком аспекте естественно не позволяет рассмотреть все гормоны организма и все их эффекты, поэтому в качестве приложения в конце пособия дается словарь, в котором представлены физиологические эффекты тех гормонов и биологически активных веществ, которые не обсуждаются в разделах пособия.
Введение Две регуляторные системы организма – нервная и гуморальная – выполняя одну функцию в организме человека и животных: приспособление к изменениям во внутренней и внешней среде, - играют свои роли по разному. Если элементарной формой нервной деятельности является рефлекс – мгновенная и точная ответная реакция организма на раздражение рецепторов, которая реализуется путем распространения нервного импульса (потенциала действия), то гуморальная регуляция осуществляется с помощью разнообразных химических веществ, которые с током крови доставляются ко всему организму. Эти вещества называются гормонами. Термин "гормон" был впервые применен в 1902 г. Старлингом и Бейлиссом в отношении открытого ими вещества, продуцирующегося в двенадцатиперстной кишке, - секретина. Термин "гормон" в переводе с греческого означает "побуждающий к действию". Гормоны - это биологически высокоактивные вещества, синтезирующиеся и выделяющиеся во внутреннюю среду организма эндокринными железами, или железами внутренней секреции, и оказывающие регулирующее влияние на функции удаленных от места их секреции органов и систем организма. Общие свойства гормонов: 1. Строгая специфичность (тропность) физиологического действия. 2. Высокая биологическая активность: гормоны оказывают свое физиологическое действие в чрезвычайно малых дозах. 3. Дистантный характер действия: клетки-мишени располагаются обычно далеко от места образования гормона. 4. Многие гормоны (стероидные и производные аминокислот) не имеют видовой специфичности. 5. Генерализованность действия. 6. Пролонгированность действия. Гормоны выполняют в организме следующие важные функции: 1. Регуляция роста, развития и дифференцировки тканей и органов, что определяет физическое, половое и умственное развитие. 2. Обеспечение поддержания гомеостаза 3. Обеспечение адаптации организма к меняющимся условиям существования. Сравнивая нервную и гуморальную формы регуляции, мы отметили, что ответ гуморальной системы наступает значительно позже, чем рефлекторная реакция. Это связано с тем, что требуется время для секреции гормона, доставки его с током крови к органам мишеням, взаимодействия с мембранными рецепторами. Гуморальная регуляция, в отличие от нервной является не локальной, а генерализованной, потому что рецепторы для гормонов, как правило, находятся в многочисленных тканях и органах. Гуморальная регуляция осуществляется более длительное время, в отличие от кратковременного рефлекторного ответа, и приводит к более выраженным изменениям в тканях, на которые оказывает эффект. В эффектах многих гормонов можно выделить: 1. срочные реакции - изменение проницаемости мембраны для ионов или глюкозы и аминокислот, что приводит, например, к сокращению гладких мышц или повышению темпа метаболизма клетки, 2. отсроченные реакции, которые заключаются в изменении активности уже существующих ферментов, и в дополнение к темпу метаболизма может измениться его направление (например, запасать глюкозу или расходовать), 3. длительные реакции - синтез новых ферментов и структурных компонентов клетки - такие реакции способны изменить и структуру, и функцию органа или системы органов. Установлены четыре основных типа физиологического действия на организм: кинетическое, или пусковое, вызывающее определенную деятельность исполнительных органов; метаболическое (изменения обмена веществ); морфогенетическое (дифференциация тканей и органов, действие на рост, стимуляция формообразовательного процесса); корригирующее (изменение интенсивности функций органов и тканей). Задумаемся над тем, что изменение метаболизма, или даже структуры органа или ткани приводит к тому, что этот орган, являясь эффектором в рефлекторной дуге, иначе реагирует на возбуждающий импульс. Таким образом, гуморальные механизмы регуляции длительно, в течение часов и суток видоизменяют при необходимости структуры, к которым будут обращены быстрые и точные нервные импульсы. Если учесть, что стимулом для выделения гормонов часто бывает возбуждающий импульс, мы получим представление о взаимодействии нервной и гуморальной регуляции, которые вместе обеспечивают надежное и эффективное приспособление организма к изменяющимся условия как внешней, так и внутренней среды организма. Коротко остановимся на взаимодействии гормонов. Каждый из гормонов или биологически активных веществ выделяется в конкретной ситуации и обладает своим спектром эффектов. Вместе с тем для каждого из гормонов существует постоянный, базовый уровень секреции. В организме человека в норме не бывает такого состояния, когда уровень любого из гормонов равен нулю. Следовательно, гормоны и БАВ могут оказывать друг на друга определенные влияния, и оказывают их. Остановимся на такой форме влияний, которую называют пермиссивные (позволяющие, обусловливающие), или сенсибилизирующие, потенцирующие эффекты. Пермиссивными называются такие эффекты, когда одни гормоны облегчают, или резко усиливают эффекты других гормонов. Например, катехоламины стимулируют гликогенолиз в печени и липолиз в адипоцитах, однако этот эффект не проявляется в отсутствии кортизола. В свою очередь катехоламины потенцируют эффекты глюкокортикоидов. Эстрогены усиливают эффекты многих гормонов, а тиреоидные гормоны увеличивают эффекты эстрогенов. Механизмы таких потенцирующих влияний неспецифичны. Если гормон или БАВ увеличивает проницаемость мембраны клетки для ионов кальция, то такая клетка будет более чувствительна к действию любого вещества. Если гормон повышает проницаемость мембраны для аминокислот и стимулирует синтез белка в клетке, то на мембране клетки увеличивается и количество мембранных рецепторов, специфичных для данной клетки, следовательно, такая клетка становится более чувствительной к действию других гормонов и БАВ. И последнее, на чем мы остановимся во введении: следует понять разницу между метаболическими или клеточными эффектами гормонов и физиологическими эффектами на уровне целого организма. Физиологический эффект – это то, к чему приводят клеточные эффекты гормонов. Например, альдостерон на уровне клеток дистальных канальцев нефрона увеличивает транспорт ионов натрия через клетки путем активации натрий/калиевого насоса, а на уровне организма такой усиленный перенос натрия реализуется в увеличении объема циркулирующей крови, увеличении минутного объема сердца и повышении системного артериального давления. Регуляция секреции гормонов (Механизмы стимуляции или подавления секреции) Говоря о стимулах для секреции гормонов, следует помнить о том, что для всех эндокринных клеток существует постоянный, базовый уровень секреции. Если нам удастся определить концентрацию гормона в крови, то мы убедимся в том, что ни для одного гормона в любой физиологической ситуации не бывает концентраций, равных нулю. Вместе с тем, концентрации гормонов, конечно, изменяются, иначе они не могли бы выполнять своих биологических функций. Что же служит стимулом для изменения темпа секреции уже синтезированного гормона? Таких стимулирующих воздействий несколько. 1. Нервный импульс, например, выделение гипоталамических либеринов и статинов, выделение адреналина и норадреналина мозговым веществом надпочечников резко возрастает при активации симпатической нервной системы. 2. Влияние другого гормона, например, тропные гормоны аденогипофиза (АКТГ, ТТГ, ФСГ и ЛГ) регулируют синтез и секрецию гормонов коры надпочечников. Щитовидной и половых желез, ангиотензин стимулирует все этапы синтеза и секрецитю альдостерона корой надпочечников. 3. Концентрация вещества, уровень которого регулируется гормоном, например концентрация глюкозы в крови служит основным стимулом для выделения инсулина островковыми клетками поджелудочной железы, высокая концентрация ионов кальция в крови непосредственно стимулирует выделение кальцитонина клетками щитовидной железы. 4. Непосредственное раздражение рецепторов (баро-, хемо-), например, стимулом для секреции АДГ в гипоталамусе может быть раздражение гипоталамических осморецепторов, а раздражение механо- и хеморецепторов кишечника приводит к секреции таких гормонов как секретин, гастрин, ХК-ПЗ. 5. Повреждение ткани, например, при травме сосуда вызывает выделение тромбоксана, простациклина; воздействие антигенов стимулирует выделение гистамина, серотонина тканями. Таким образом, для одного гормона возможно несколько вариантов стимуляции и подавления секреции. Такая организация эндокринной регуляции делает ее надежной, и способной обеспечить необходимую перестройку работы организма в любой ситуации. Транспорт Прежде, чем обсуждать варианты транспорта гормонов, вспомним о том, что есть гормоны, которые оказывают свои эффекты непосредственно в том месте, где секретируются, например, гастрин в слизистой желудка. Такое действие называется паракринным. Итак, возможны следующие варианты транспорта гормонов: 1. Транспорта нет – паракринный эффект. 2. Транспорт со специальным белком, например, таким образом транспортируются глюкокортикоиды со специальным белком – транскортином. В состоянии покоя 80% концентрации таких гормонов приходится на комплекс со специфическими белками. Биологическая активность определяется содержанием свободных форм гормонов. Связанные формы гормонов являются как бы депо, физиологическим резервом, из которого гормоны переходят в активную свободную форму по мере необходимости. 3. Транспорт в неактивной или малоактивной форме, например, щитовидная железа выделяет два гормона: трииодтиронин и тироксин. Тироксин – менее активная форма, в этом виде гормон переносится в крови, а в клетках, после дейодирования, превращается в активную форму – трииодтиронин. 4. Транспорт в свободном виде, таким образом транспортируются преимущественно гормоны белковой природы. Рисунок 4. Механизм действия стероидных гормонов Для реализации эффекта стероидных гормонов необходим внутриклеточный (а для некоторых гормонов внутриядерный) рецептор, который не только перемещает гормон к хромосомам, но и определяет взаимодействие в определенном месте и экспрессирование генома для синтеза информационной РНК. Физиологические эффекты Следует отличать физиологические эффекты гормонов от механизмов их действия: механизм действия – это то, что происходит в клетках под действием гормонов, физиологический эффект - те изменения, которые при этом происходят на уровне организма. Например, альдостерон увеличивает реабсорбцию натрия в дистальных канальцах нефронов, а на уровне организма задержка натрия приводит к увеличению объема циркулирующей крови и повышению АД. Синтез структурных компонентов клетки под действием соматотропного гормона приводит к росту и развитию всего организма. Метаболизм и выведение Инактивация гормонов происходит в эффекторных органах в результате воздействия ферментов. Большая часть гормонов инактивируется в печени, где гормоны претерпевают различные химические изменения путем связывания с глюкуроновой или серной кислотой. Частично гормоны выделяются с мочой в неизмененном виде. Действие некоторых гормонов может блокироваться благодаря секреции гормонов, обладающих антагонистическим эффектом.
Вопросы и задания для самоконтроля 1. Перечислите несколько стимулирующих факторов, которые увеличивают секрецию гормонов, приведите примеры. 2. Опишите механизм действия белково-пептидных гормонов, назовите вторичные посредники. 3. Опишите механизм действия стероидных гормонов, объясните почему эффект стероидных гормонов наступает значительно позднее эффекта пептидных гормонов. 4. Перечислите формы транспорта гормонов в крови. 5. Объясните разницу между паракринными и эндокринными эффектами. 6. Объясните, каким образом происходит удаление гормонов из крови.
Рисунок 5. Гипоталамический нейрон
Каким же образом рилизинг-гормоны включаются в систему гуморальной регуляции деятельности организма? Гипоталамус, который является центром регуляции вегетативных функций, как известно, конвергирует на своих нейронах огромное количество информации. Эти информационные потоки условно можно разделить на несколько групп: а) информация, поступающая по восходящим спинномозговым путям (в основном температурной и болевой чувствительности) от всего организма; б) информация, поступающая по чувствительным веточкам черепно-мозговых нервов – это информация от сердца, сосудов, дыхательной, пищеварительной систем, лица; в) информация, поступающая от органов чувств; г) информация от лимбической системы, которая организует эмоциональную реакцию организма и от коры больших полушарий; д) информация, поступающая не нервным, а гуморальным путем (кровь, мозговая жидкость) о содержании в крови глюкозы, аминокислот, ее осмотической концентрации, температуре, о содержании в крови гормонов. Этот информационный поток обрабатывается ЦНС, приводит к осуществлению определенных безусловных рефлексов, вызывает какие-то изменения поведения и, наряду с этим, стимулирует выделение нейронами гипоталамуса рилизинг-гормонов. Необходимо уточнить, что те из них, которые стимулируют секрецию гормонов клетками гипофиза, называются либерины, а подавляющие секрецию – статины. Рилизинг гормоны, которые с током крови доставляются в аденогипофиз, изменяют темп секреции гормонов аденогипофиза. Эти гормоны, в свою очередь, оказывают влияние на периферические эндокринные железы и ткани организма (рисунок 6).
Рисунок 6. Рисунок 7. Гормоны гипофиза Между рилизинг-гормонами и аденогипофизарными гормонами существует не только система прямых связей, но и обратная связь. Эта связь проявляется в том, что аденогипофизарные гормоны, выделившись в кровь, тормозят секрецию рилизинг-гормонов гипоталамуса. Гормоны аденогипофиза можно объединить в семейства по их химическому строению. Обратите внимание на то, что подобные аденогипофизарным гормоны синтезируются и в плаценте.
Теперь, когда мы знаем названия аденогипофизарных гормонов, будет легко каждому предоставить свой рилизинг-гормон.
Либерины и статины гипоталамуса, действующие на аденогипофиз.
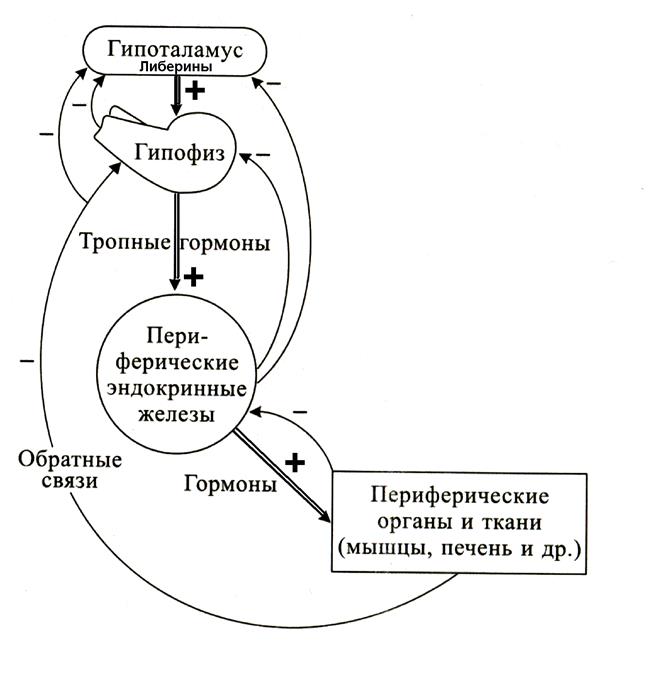
Итак, гипоталамические либерины и статины регулируют секреторную деятельность аденогипофиза. Гормоны аденогипофиза оказывают влияние и на железы внутренней секреции, и на ткани организма. В результате повышается секреторная активность желез, находящихся под управлением гипофиза, и метаболизм в тканях. В периферической крови увеличивается уровень гормонов периферических желез и продуктов метаболизма тканей. Эта информация с током крови достигает и гипофиза, и гипоталамуса и приводит к снижению секреторной активности. Таким образом, в гипоталамо-гипофизарной системе работают два варианта обратных связей, в которых информация передается в виде концентрации гормонов: между гипофизом и гипоталамусом короткие обратные связи, а между периферическими железами и гипоталамусом – длинные (рисунок 8). Нейроны гипоталамуса организуют эндокринные функции трех основных гормональных систем:
Рисунок 8. Прямые и обратные связи в гипоталамо-гипофизарной системе Вопросы и задания 1. Нарисуйте схему связей гипоталамуса и гипофиза. 2. Перечислите гипоталамические либерины и основные стимулы, приводящие к увеличению их секреции. 3. Объясните, каким образом либерины поступают к аденогипофизу. 4. Перечислите тропные гормоны аденогипофиза. 5. Перечислите компоненты гипоталамо-гипофизарно надпочечниковой системы, нарисуйте схему прямых и обратных связей 6. Перечислите компоненты гипоталамо-гипофизарно тиреоидной системы, нарисуйте схему прямых и обратных связей. Половые гормоны Пол человека, как известно, определяется сочетанием половых хромосом: две Х хромосомы у млекопитающих и человека определяют женский, а Х и Y - мужской пол. Генетическая детерминация пола будущего организма происходит при оплодотворении. Однако, для реализации генетической программы пола необходимо влияние половых гормонов, которые синтезируются в половых железах. Половые железы, или гонады - семенники (яички) у мужчин и яичники у женщин относятся к числу желез со смешанной секрецией. Внешняя секреция связана с образованием мужских и женских половых клеток - сперматозоидов и яйцеклеток. Эндокринная функция заключается в секреции мужских и женских половых гормонов и их выделении в кровь. Как семенники, так и яичники синтезируют и мужские и женские половые гормоны, но у мужчин преобладают андрогены, а у женщин - эстрогены. Половые гормоны способствуют эмбриональной дифференцировке, в последующем развитию половых органов и появлению вторичных половых признаков, определяют половое созревание и поведение человека. В женском организме половые гормоны регулируют овариально-менструальный цикл, а также обеспечивают нормальное протекание беременности и подготовку молочных желез к секреции молока. Мужские половые гормоны Хромосома Y содержит специфические гены, располагающиеся в детерминирующей пол области SDR (от Sex-Determinig Region — область детерминации пола). Один из таких генов кодирует фактор TDF (от Testis-Determining Factor), определяющий развитие мужских гонад. Мутации гена, кодирующего TDF, приводят к реверсии (обращению) пола и дисгенезии (неправильному развитию) гонад, а генетические мужчины (46XY) имеют женский фенотип. Критическая стадия развития гонад — 8‑я неделя внутриутробного развития. До 45–50 дня зачатки гонад не имеют половой дифференцировки. Под влиянием регуляторного фактора TDF развиваются яички; при отсутствии этого фактора развиваются яичники. В это время в яичках дифференцируются клетки Лейдига, которые под контролем гонадотропинов (хорионического и гипофизарного) секретируют тестостерон. Экспрессия TDF в клетках Сертоли яичек инициирует транскрипцию гена, кодирующего МИФ (мюллеров ингибирующий фактор, или фактор регрессии мюллеровых каналов). Дальнейшую дифференцировку других структур мужской половой системы определяют мужские половые гормоны и пептидный гормон МИФ, продуцируемые в яичках плода. Итак, в эмбриогенезе осуществляется синтез тестостерона, синтез рецепторов с нему, причем не только в гонадах, но и во всем организме, синтез редуктазы в тканях (фермент, преобразующий Т в 5a ДТ), синтез фактора регрессии Мюллеровых каналов. Таким образом, к моменту рождения сделан первый шаг в реализации генетической программы пола, но реализация не завершена. На данном этапе: 1) определен тип гонад – мужской, 2) реализована гормональная программа в соответствии с полом – синтез тестостерона, 3) сформирован механизм приема этой гормональной программы – рецепторы к тестостерону во всем организме плюс ферментативная активность редуктазы. Вторая волна реализации генетической программы пола наблюдается в пубертатный период. Обратим внимание на важную особенность тестостерона: существуют два типа эффектов детерминирующие, которые определяют необратимые структурные изменения и регулирующие, которые проявляются в течение всего репродуктивного периода жизни мужчины. В этот период в организме мальчика происходит окончательное формирование и дозревание мужской половой системы, появление просвета в канальцах яичек. Окончательное подавление роста молочных желез. В головном мозге регистрируется окончательное подавление деятельности циклического центра гипоталамуса (начало положено в первую волну), эту роль в основном выполняют эстрогены. Формируется вариант секреции СТГ по мужскому типу – импульсная активность с высокими волнами, мужской вариант поведения с более выраженной, по сравнению с женским, агрессивностью. В печени заканчивается формирование типа метаболизма по мужскому типу. Для этого варианта характерен «расходный» тип метаболизма, более интенсивный синтез белков, необходимых для обеспечения роста и работы репродуктивной системы, высокая реактивность в процессе адаптации. В женском организме «накопительный» вариант метаболизма обеспечивает резервирование энергетических и структурных запасов, это настройка на накопление к периоду вынашивания беременности. Реализация генетической программы пола завещается окончательным формированием вторичных половых признаков, к которым относится и форма тела, обусловленная более мощным скелетом, мышечной массой и характером отложения жира, и тип распределения волос по телу, и тембр голоса, и психофизиологические черты. К завершению пубертатного периода генетическая программа пола реализована. Андрогены синтезируются группами специализированных клеток, лежащих в интерстиции между извитыми семенными канальцами. Эти клетки называют клетками Лейдига. Известно несколько стероидных гормонов (тестостерон, дигидротестостерон, дегидроэпиандростерон, андростендион и некоторые другие) с андрогенной активностью. Тестостерон основной циркулирующий андроген, суточная секреция — 5 мг (от 2 мг до 10 мг), синтезируется в клетках Лейдига. Гормон удаляется из крови в течение 30–60 минут, присоединяясь к клеткам–мишеням или распадаясь на неактивные компоненты. Ароматизация тестостерона ведёт к образованию эстрадиола. Транспортные белки: андрогенсвязывающий белок отвечает за поддержание высокого уровня тестостерона в сперматогенном эпителии путём накопления тестостерона в просвете семенных канальцев. b-глобулин и альбумин связывают в крови до 99% тестостерона. Рецептор андрогенов (рецептор дигидротестостерона) относится к ядерным, он содержит ДНК-связывающую область. Тестостерон свободно проникает в цитоплазму клеток–мишеней. Под влиянием 5a-кеторедуктазы тестостерон превращается в дигидротестостерон, который связывается рецепторным цитоплазматическим белком. Этот комплекс мигрирует в клеточное ядро и активирует экспрессирование ряда генов и транскрипцию. Итак, тестостерон участвует в созревании мужских половых клеток - сперматозоидов, которые образуются в сперматогенных эпителиальных клетках семенных канальцев. Тестостерон обладает выраженным анаболическим действием, т.е. увеличивает синтез белка, особенно в мышцах, что приводит к увеличению мышечной массы, к ускорению процессов роста и физического развития. За счет ускорения образования белковой матрицы кости, а также отложения в ней солей кальция гормон обеспечивает рост, толщину и прочность кости. Способствуя окостенению эпифизарных хрящей, половые гормоны практически останавливают рост костей в длину, это происходит в конце пубертатного периода. Тестостерон уменьшает содержание жира в организме. Гормон стимулирует эритропоэз, чем объясняется большее количество эритроцитов у мужчин, чем у женщин. Тестостерон оказывает влияние на деятельность центральной нервной системы, определяя половое поведение и типичные психофизиологические черты мужчин. Рисунок 9. Регуляция секреции и эффекты тестостерона
Женские половые гормоны К женским половым гормонам относятся эстрогены и прогестерон. В фолликулах яичников осуществляется синтез эстрогенов, желтое тело яичника продуцирует прогестерон. Основная функция по регуляции роста организма, его развития ы соответствии с полом, репродуктивной функции принадлежит эстрогенам. К эстрогенам относятся: Е1 –эстрон, Е2 – эстрадиол, Е3 – эстриол, Е4 – эстетрон, основной эстроген - Е2. Для формирования женской половой системы в эмбриогенезе не нужно специальной программы, поэтому если нет фактора регрессии мюллеровых каналов, развитие организма идет по женскому типу, при этом вырабатываются почти одинаковые количества эстрогенов и андрогенов. В пубертатном периоде эстрогены, прежде всего, завершают формирование органов женской половой сферы: влагалища, матки, фаллопиевых труб. Эстрогены стимулируют развитие и рост молочных желез. Одним из важнейших эффектов эстрогенов является их сенсибилизирующий эффект по отношению ко многим гормонам: прогестерону, пролактину, андрогенам, катехоламинам. Под влиянием эстрогенов усиливается синтез белка в печени, синтезируются транспортные альбумины для переноса витаминов и гормонов, анитигены, липопротеины, ферменты, метаболизирующие глюкокортикоиды. В гипоталамусе эстрогены стимулируют циклические центры, оказывают непосредственное действие на систему гипоталамус – гипофиз – гонады, активируют синтез окситоцина и вазопрессина. Кроме этого эстрогены влияют на развитие костного скелета, ускоряя его созревание. За счет действия на эпифизарные хрящи они тормозят рост костей в длину. Эстрогены оказывают выраженный анаболический эффект, усиливают образование жира и его распределение, типичное для женской фигуры, а также способствуют оволосению по женскому типу. Эстрогены задерживают азот, воду, соли. Под влиянием этих гормонов изменяется эмоциональное и психическое состояние женщин. Во время беременности эстрогены способствуют росту мышечной ткани матки, эффективному маточно-плацентарному кровообращению, вместе с прогестероном и пролактином - развитию молочных желез. Эффекты эстрогенов
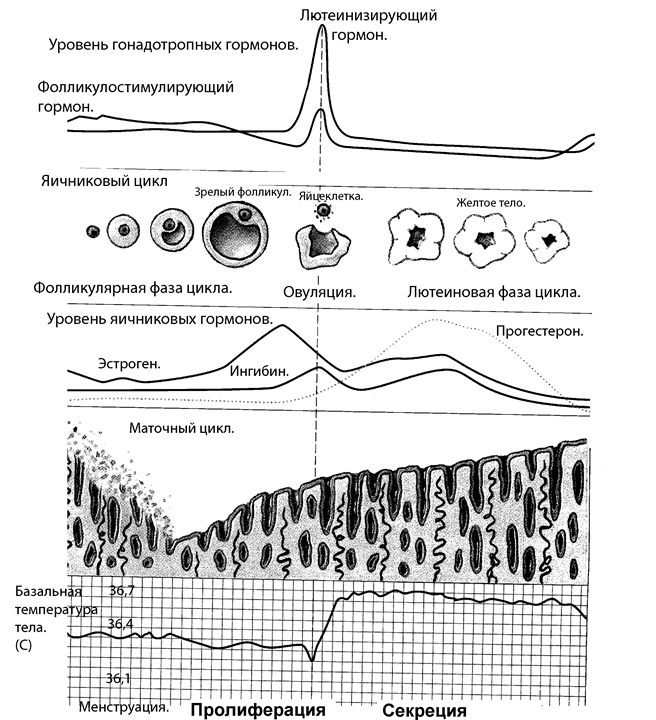
Фолликулиновая фаза цикла завершается овуляцией, которая происходит на пике секреции ЛГ, обратим внимание на то, что без лютеинизирующего гормона овуляции не происходит, в этом случае цикл становится ановуляторным. После овуляции в желтом теле яичника, которое развивается на месте лопнувшего фолликула, начинает вырабатываться гормон - прогестерон. Регуляция секреции прогестерона обеспечивается ЛГ. Главная функция прогестерона - подготовка эндометрия к имплантации оплодотворенной яйцеклетки и обеспечение нормального протекания беременности. Если оплодотворение не наступает, желтое тело дегенерирует. Прогестерон вместе с эстрогенами регулирует морфологические перестройки в матке и молочных железах, усиливая процессы пролиферации и секреторной активности. В результате этого в секрете желез эндометрия возрастают концентрации липидов и гликогена, необходимых для развития эмбриона. Прогестерон усиливает гиперплазию и гипертрофию миометрия во время беременности и снижает возбудимость матки. У небеременных женщин прогестерон участвует в регуляции менструального цикла. Гормон способствует росту альвеол в молочных железах. В гипоталамусе высокие концентрации прогестерона подавляют секрецию всех тропных гормонов: ФСГ, ЛГ, ПРл. Именно поэтому и происходит кровотечение – отторжение нового слизистого слоя с сосудами. В головном мозге под влиянием прогестерона наблюдается снижение возбудимости. Прогестерон усиливает основной обмен и повышает базальную температуру тела, что используется в практике для определения времени наступления овуляции. Регуляция менструального цикла осуществляется гонадолиберином и гонадотропинами. ГОНАДОЛИБЕРИН -гипоталамический декапептид, который регулирует секрецию и фолликулостимулирующего (ФСГ) и лютеинизирующего (ЛГ) гормонов. Способность ГЛ в одних случаях стимулировать синтез и высвобождение ФСГ, а в других ЛГ объясняется модулирующим влиянием половых гормонов (различный уровень во время полового цикла) и участием простагландинов. ГОНАДОТРОПИНЫ -гормоны аденогипофиза - фолликулостимулирующий (ФСГ) и лютеинизирующий (ЛГ). Рисунок 10. Регуляция секреции и основные эффекты Женских половых гормонов
Рисунок 11. Овариально-менструальный цикл
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 549; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.9.183 (0.013 с.) |