Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Тема 8: Сцепленное наследование. Генетические карты.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Тема 9: Генетика пола
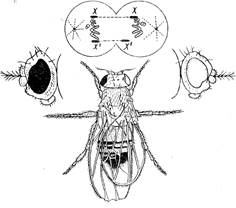
Пол – это совокупность признаков и свойств организма, обеспечивающих его участие в воспроизводстве потомства и передаче наследственной информации за счет образования гамет. Обычно признаки, по которым отличаются особи разных полов, делят на первичные и вторичные половые признаки. К первичным относят те морфологические и физиологические особенности организма, которые обеспечивают образование гамет и соединение их в процессе оплодотворения. К числу их относятся, например, гонады, половые пути и наружные гениталии у высших животных, андроцей и гинецей у высших растений. К вторичным половым признакам относят признаки и свойства организма, непосредственно не обеспечивающие процессы гаметогенеза, спаривания и оплодотворения, но играющие некоторую вспомогательную роль в половом размножении. К ним относятся особенности строения плавников у рыб, оперения у птиц, грудных желез у млекопитающих, длины междоузлий и сроков цветения у высших растений и др. Из числа вторично-половых признаков выделяют иногда ограниченные полом признаки. Генетическую информацию о них несут все особи, но проявляются они только у одного пола. Так, быки несут гены, определяющие молочность, петухи - гены, определяющие яйценоскость, но действие их у самцов не проявляется. Следует отличать также признаки, зависимые от пола. К числу зависимых от пола признаков относятся такие признаки, характер доминирования которых в гетерозиготе зависит от пола особи. Так, доминантные гомозиготные овцы (НН) рогаты, а гомозиготные рецессивы (hh) безроги, независимо от пола. Однако гетерозиготные (Hh) самцы рогатые, а самки безрогие. Аналогично наследуется раннее облысение у человека. Доминирование в таких случаях определяется количеством мужских и женских половых гормонов в крови. Знание закономерности наследования признаков, зависимых от пола, позволяет еще глубже понять сущность явления доминирования и подойти к практическому управлению доминированием. Различия особей мужского и женского пола - половой диморфизм встречается у некоторых низших и многих высших растений и животных. Среди высших растений около 5000 видов двудомны. У высших животных обоеполые (гермафродитные) формы в норме не встречаются. Во всех случаях отличия касаются морфологических, биохимических, физиологических признаков, экологии, поведения у животных и т. д. Однако на низших ступенях эволюции у одноклеточных организмов отличия особей, вступающих в копуляцию, могут быть невидимы и касаться лишь физиологических особенностей, поэтому и пол их обозначают условно как «+ пол» или «-пол». Различия между этими двумя типами часто носят количественный характер, так что у многих одноклеточных водорослей (хламидомонады, например), некоторых низших грибов или инфузорий удается выделить несколько линий, внутри которых копуляция клеток невозможна, а между линиями она осуществляется. Удалось сгруппировать многочисленные линии в «+» и «-» типы, но оказалось, что и внутри типа копуляция клеток разных линий иногда происходит. Следовательно, здесь можно говорить об относительной сексуальности. Относительная сексуальность особенно хорошо изучена у хламидомонад М. Гартманом и Ф. Мевусом. Они показали, что гаметы даже одного пола, но с разной половой валентностью способны воспроизводить потомство половым путем. Половые различия гамет связаны с гормоноподобными веществами (гамонами). Научное обоснование полового диморфизма стало возможным благодаря развитию цитологии и генетики. У всех раздельнополых организмов были обнаружены хромосомы, одинаковые у обоих полов, и хромосомы, по которым, мужской и женский пол отличаются друг от друга. Хромосомы одинаковые для женских и мужских особей назвали аутосомами, а отличающиеся- половыми, или гетерохромосомами (гоносомами). Например, в клетке человека содержится 46 хромосом или 23 пары: 22 пары аутосом и 1 пара половых хромосом. Соотношение полов у раздельнополых организмов близко 1: 1, т.е. самцы и самки встречаются одинаково часто. Еще Мендель обратил внимание, что такое расщепление характерно для скрещивания особей: Аа х аа. Из это следует, что один из полов должен быть гомозиготным, а другой – гетерозиготным. Пол, образующий одинаковые в отношении определения пола гаметы, называют гомогаметным, а пол, образующий разные гаметы – гетерогаметным. В случае гетерогаметности мужских особей половые хромосомы обозначают как X- и Y-хромосомы. В случае гетерогаметности женских особей самка обозначается ZW, самец – ZZ. Изучение половых хромосом показало, что они отличаются от аутосом и генетически и цитологически. В них содержится меньшее число аллелей, они богаты гетерохроматином, редупликация их происходит асинхронно с аутосомами, а у гомогаметного пола одна из Х хромосом репродуцируется позже остальных. В мейозе гоносомы часто спирализованы (гетеропиктоз). Половые хромосомы у гетерогаметного пола не конъюгируют или конъюгируют лишь частично, что указывает на частичную гомологичность (гемизиготность). В этом случае проявляется рецессивный ген, имеющийся в генотипе в единственном числе. Так X-хромосома может содержать ген, вызывающий гемофилию (пониженную свертываемость крови). Тогда все мужские особи, получившие эту хромосому, будут страдать этим заболеванием, так как Y-хромосома не содержит доминантного аллеля. Анализ половых хромосом у различных организмов позволил сформулировать хромосомную теорию определения пола и показал, что существует 5 типов хромосомного определения пола: 1) ♀ XX, ♂ ХУ - характерен для млекопитающих (в том числе и для человека), червей, ракообразных, большинства насекомых (в том числе для дрозофил), большинства земноводных, некоторых рыб (рис. 4). 2) ♀ ZW, ♂ ZZ - характерен для птиц, пресмыкающихся, некоторых земноводных и рыб, некоторых насекомых (чешуекрылые). 3) ♀ XX, ♂ Х0 - (0 обозначает отсутствие хромосом) встречается у некоторых насекомых (прямокрылые). 4) ♀ Z0, ♂ ZZ - встречается у некоторых насекомых (равнокрылые). 5) гапло-диплоидный тип (♀ 2n, ♂ n) встречается, например, у пчел и муравьев: самцы развиваются из неоплодотворенных гаплоидных яйцеклеток (партеногенез), самки - из оплодотворенных диплоидных.
Рисунок 4. Схема определения пола у дрозофилы. Хромосомные наборы: 1 - соматических клеток; 2-— сперматозоидов; 3 - яйцеклеток; 4 - потомства.
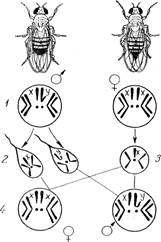
Таким образом, хромосомный механизм определения пола является идеальным саморегулирующимся механизмом. В результате оплодотворения возникает равное количество самцов и самок. Однако, иногда встречаются такие явления, которые как будто специально созданы природой для проверки правильности теории. В отношении хромосомной теории примером может служить явление гинандроморфизма. Организмы, совмещающие в себе части тела разных полов - мужского и женского, называют гинандроморфами (гин -♀, андр - ♂). Гинандроморфы существуют у тех видов, у которых четко выражен половой диморфизм (насекомые, птицы, человек), но встречаются они редко. При латеральном гинандроморфизме, например у дрозофилы, одна половина тела имеет признаки женского пола, а другая - мужского (рис. 5). Как может возникнуть такой организм? Цитологические исследования показывают, что ткани гинандроморфа химерны: женская половина несет две Х-хромосомы, а мужская - одну. На приведенном рисунке показан случай, когда у гинандроморфа рецессивный, сцепленный с полом ген white проявился на мужской стороне тела и не проявился на женской. Почему это так? У гинандроморфа, возникшего из зиготы w+w, при первом делении дробления в силу каких-то необычных условий одна из Х-хромосом, несущая ген w+, в одной из дочерних клеток (бластомеров) утрачивается. Тогда две дочерние клетки окажутся неодинаковыми в отношении Х-хромосом: одна w, а вторая w. w+
Рисунок 5. Гинандроморф дрозофилы. Вверху - предполагаемая схема возникновения; X - элиминируется.
Половина тела мухи, развившаяся из первой клетки, окажется женской и с красным глазом, а из второй разовьется половина тела с признаками мужского пола и с белым глазом, поскольку рецессивный ген w, содержащийся в единственной Х-хромосоме, будет в гемизиготном состоянии. Таким образом, и цитологический, и генетический анализ показывает, что в данном случае причиной гинандроморфизма может быть элиминация одной из Х-хромосом. Кроме этого типа гинандроморфизма, который можно назвать монозиготным, известен также дизиготический гинандроморфизм. Он обнаружен у бабочек - Abraxas, тутового шелкопряда и у дрозофилы. Например, иногда в яйцеклетке тутового шелкопряда (самка гетерогаметна) образуются два женских пронуклеуса, один из которых кроме аутосом (обозначим их А) содержит Х-хромосому (Х + А), а другой — У + А. При полиспермии оба пронуклеуса будут оплодотворены разными спермиями, тогда в одном из бластомеров будет ХХ + АА, а в другом — ХУ+АА. Это и приведет к развитию дизиготного гинандроморфа. Аналогично может возникать гинандроморф у дрозофилы, только здесь различия между бластомерами получаются за счет разных сперматозоидов (самцы гетерогаметны). Исключения из хромосомной теории определения пола. По мере накопления фактов хромосомная теория определения пола не только находила подтверждение, но и встречала некоторые трудности. Оставался открытым вопрос о том, не являются ли половые хромосомы индикаторами пола, вторичнополовыми признаками? Анализ исключительных особей у дрозофил, которые были получены в опытах Бриджеса, как результат нерасхождения половых хромосом показал, что особи, имеющие, кроме аутосом, ХХУ-хромосомы (ХХУ + АА), являются самками, а особи ХО + АА—самцами. Эти факты убедительно говорили о том, что половые хромосомы отнюдь не индикаторы пола. Но как же они определяют пол, если особи ХУ+АА и ХО+АА являются самцами, а ХХ+АА и ХХУ+АА самками? Очевидно, дело обстоит не так просто, как это казалось вначале. Балансовая теория определения пола – гипотеза Бриджеса. К. Бриджес обнаружил у дрозофилы несколько самок, имевших триплоидный набор хромосом 3Х + 3А. При скрещивании их с диплоидными самцами ХУ + 2А было получено потомство, морфологическое, цитологическое и генетическое обследование которого выявило восемь типов особей с различным соотношением половых хромосом и наборов аутосом: 3Х: 3А; 2Х: 2А; (2Х + У): 2А; 2Х: 3А; (2Х + У): 3А; ХУ: 2А; 3Х:2А; ХУ: А. Появление анеуплоидных мух обусловлено нарушением нормальной конъюгации и расхождения хромосом в мейозе у триплоидных самок. Преобладание наборов аутосом в женской зиготе (2Х: 3А) привело к образованию особей с промежуточным развитием пола, т. е. интерсексов. Уменьшение числа Х-хромосом (X: 2А) привело к развитию мужского пола. Но если число наборов аутосом увеличивается до трех при наличии одной Х-хромосомы (Х:3А), развивается сверхсамец - организм с гипертрофированными признаками самца. Он оказывается стерильным. Напротив, увеличение числа Х-хромосом при диплоидном числе аутосом (3Х:2А) ведет к развитию сверхсамки с ненормально развитыми яичниками и с другими нарушениями признаков пола. На основании этих опытов К. Бриджес пришел к выводу, что у дрозофилы женский пол определяет не присутствие двух Х-хромосом, а мужской - ХУ, а соотношение числа половых хромосом и наборов аутосом является решающим в определении пола особи. Гены женской тенденции сосредоточены главным образом в Х-хромосомах, гены мужской - в аутосомах. Это видно из того, что все особи с балансом хромосом (или половым индексом) Х:А = 1 представляют собой самок, отношение X: 2А = 0,5 дает самцов; баланс хромосом с отношением от 1 до 0,5 определяет интерсексуальность. Отношение трех Х-хромосом к двум наборам аутосом 3Х: 2А = 1,5 ведет к развитию сверхсамок. Напротив, увеличение количества наборов аутосом на одну Х-хромосому X: 3А = 0,33 определяет развитие сверхсамцов. Эти формы называют суперсексами, У-хромосома у дрозофилы пол не определяет. В таблице 1 приведены различные половые типы и соответствующие им половые индексы у дрозофилы (по Бриджесу). Таблица 1 Определение пола у дрозофилы согласно балансовой теории
Примечание. Звездочкой отмечены не встречающиеся гаплоидные особи, однако участки тканей такой структуры наблюдались в диплоидных особях. Эти участки обнаруживали женские признаки.
Для доказательства того, что пол у дрозофилы определяется балансом многих генов, а Х-хромосома определяет женский пол и аутосомы - мужской, были проведены дополнительные остроумные опыты. Методом хромосомных перестроек (дупликаций) стали получать особей, имеющих, кроме двух Х-хромосом и трех наборов аутосом, дополнительные участки Х-хромосомы различной длины. По мере удлинения таких участков интерсексы становились все более похожими на самок, как и следовало ожидать. Кроме того, в потомстве триплоидных самок в ряду поколений была проведена селекция на проявление интерсексуальности, т. е. на способность давать интерсексов либо женского, либо мужского типа. При этом соотношение хромосом у триплоидных самок этих двух линий сохранялось прежним — 3Х: 3А, так же как и соотношение хромосом у интерсексов, а проявление интерсексуальности у потомства изменялось в соответствии с направлением селекции. Гипотеза Гольдшмидта. Еще в 1911 г. Р. Гольдшмидт показал, что при реципрокных скрещиваниях различных географических рас непарного шелкопряда (Limantria dispar) в потомстве наряду с самцами и самками возникают формы, имеющие постепенные переходы в отношении первичных и вторичных половых признаков от мужского пола к женскому, т. е. интерсексы. Интерсексы имели хромосомный набор, типичный или для самок, или для самцов. Гольдшмидт объяснил полученные результаты следующим образом. У бабочек непарного шелкопряда гетерогаметным полом (ХУ) является женский, а гомогаметным (XX) - мужской. Можно представить, что интерсексуальность в этом случае объясняется балансом двух факторов: женского (в У-хромосоме) и мужского (в Х-хромосоме). При этом играет роль «сила» и «слабость» этих факторов у разных рас, а не соотношение хромосом, как это имело место у дрозофилы. Если скрещивается самка «слабой» расы с самцом «сильной» расы, то все самцы будут нормальными, а самки - интерсексами. В этом случае гены, определяющие мужской пол, локализованные в Х-хромосоме, пришедшей от самца, «сильнее», чем гены, определяющие женский пол в У-хромосоме, пришедшей от «слабой» матери. Следовательно, и эта гипотеза определения пола также может быть названа гипотезой «генного баланса». Универсальность балансовой теории определения пола. Многочисленные факты, накопленные генетикой в последние годы, позволили сформулировать балансовую теорию определения пола, согласно которой пол особи определяется балансом генов, детерминирующих мужской и женский пол и локализованных в любых хромосомах генома. Так, у человека, в отличие от дрозофилы, У-хромосома играет большую роль в определении пола. При отсутствии У-хромосомы и любом числе Х-хромосом особь фенотипически определяется как женская. Наличие У-хромосомы определяет развитие мужского пола. У тутового шелкопряда У-хромосома сдвигает баланс, определяя женский пол, независимо от числа Х-хромосом в кариотипе и степени плоидности. Балансовая теория определения пола оказалась приложимой и к высшим растениям. У дремы (Melandrium) пол определяется прежде всего балансом Х- и У-хромосом. Изменение количества аутосом эффекта не дает. Однако даже женские растения, лишенные У-хромосомы, имеют мужские потенции. Так, при поражении женских растений головней в цветках развиваются тычинки, т. е. цветки становятся морфологически гермафродитными. Однако известны случаи, когда пол особей определяется не балансом хромосом, а взаимодействием нескольких генов. Так, у пчел, например, из оплодотворенных яиц развиваются диплоидные самки, а из неоплодотворенных (партеногенетически) - самцы. Диплоидия соматических тканей восстанавливается лишь в процессе развития самца. Однако можно предполагать, что не сам факт оплодотворения определяет пол у пчел, как это считали до сих пор. Так, у наездника (Habrobracon), близкого к пчеле вида, из оплодотворенных яиц развиваются самки, а из неоплодотворенных - самцы. Но описаны случаи, когда при близкородственном спаривании из оплодотворенных яиц развиваются самцы. Генетический анализ показал, что женский пол у наездника определяется комплементарно гетерозиготным состоянием двух аллелей, независимо от наличия или отсутствия оплодотворения: ха х+ , а мужской – гомозиготным ха х+ или x+ xb х+ хв ха х+ x+ xb Сейчас получены данные, что и у пчел из оплодотворенных яиц могут иногда выводиться трутни. Может быть, и у них имеет место аналогичный механизм взаимодействия генов. Балансовая теория определения пола сейчас является общепринятой. Она показывает генетически обусловленную потенциальную бисексуальность всех раздельнополых организмов и их гамет. При этом, механизмы, поддерживающие баланс генов, могут быть разными.
Дифференциация пола Вслед за определением пола следует дифференциация, т. е. развитие половых различий: формирование воспроизводительной системы, а также физиологического и биохимического механизмов, обеспечивающих скрещивание. Зачаточные индифферентные в половом отношении гонады у эмбрионов животных имеют двойственную природу. Они состоят из внешнего слоя - кортекса (cortex), из которого в процессе дифференциации развиваются женские половые клетки, и из внутреннего слоя - медуллы (medulla), из которого развиваются мужские гаметы (рис. 6). В ходе дифференциации пола идет развитие одного из слоев гонады и подавление другого. У мужского пола быстрее развивается медуллярная ткань, которая подавляет деятельность кортикального слоя, в результате гонады превращаются в семенники. У женского пола ускоряется развитие кортикального слоя, в силу чего подавляется формирование медуллярного слоя и гонады превращаются в яичники. В соответствии с этими преобразованиями дифференцируются и половые пути, которые тоже закладываются одинаковыми у особей обоих полов. Процесс дифференциации пола у многих животных обусловлен гормонами, которые выделяются не только эндокринными железами, но кортикальным и медуллярным слоями полового зачатка, а в последующем - и половыми железами. Эти вещества отличаются по характеру своего действия и по времени их выработки. У особей мужского пола преобладает «мужское вещество» - медулларин, у женского - кортикальный гормон.
Рисунок 6. Схема дифференциации гонад в онтогенезе.
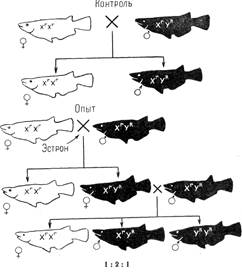
Дальнейшая дифференциация пола, особенно развитие вторичных половых признаков, также идет под влиянием различных гормонов. Уровень гормональной секреции контролируется генами, точнее, их балансом. Преобладание генов, определяющих мужской пол, в общем балансе приводит к повышению активности мужских гормонов и к дифференциации мужского пола, обратное соотношение генов - к развитию женского пола. Смена активности гормональной секреции то одного, то другого пола в онтогенезе приводит к развитию интерсексуальных форм. На дифференциацию пола у высших растений значительное влияние оказывают растительные гормоны - ауксины. У многих форм организмов в процессе онтогенеза может происходить переопределение пола в естественных или искусственных условиях. Это явление служит наилучшим доказательством наследственной бисексуальности. У млекопитающих при развитии разнополых близнецов иногда происходит изменение пола одного из них в эмбриогенезе. Так, у разнополых двоен крупного рогатого скота бычки развиваются нормально, а телочки часто оказываются интерсексами. Такие животные были названы фримартинами; они, как правило, бесплодны. Подобные изменения вызываются тем, что семенники раньше начинают выделять мужские гормоны в кровь, чем яичники. Этот пример не единичен. Известны и другие случаи переопределения пола в онтогенезе особи без специального воздействия. Однако в последние годы все больше привлекает внимание исследователей экспериментальное переопределение пола. Один из замечательных примеров полного переопределения пола в онтогенезе получен на аквариумных рыбках медаки (Oryzias latipes), у которых доминантный ген красной окраски R находится в У-хромосоме, а его рецессивная аллель r – в Х-хромосоме. В этом случае самцы (ХrУR) всегда будут красными, а самки, имеющие генотип XrXR, - белыми. При изучении наследования этого признака в нескольких поколениях, скрещивание ♀ХrХr и ♂ ХrУR неизменно давало белых самок и красных самцов (рис. 7). В опыте выклюнувшиеся, еще не дифференцированные в половом отношении мальки получали в корме в течение 8 месяцев добавку женского полового гормона (эстрона или стилбестрола). В результате оказалось, что все рыбки (белые и красные) по фенотипу были самками, с нормальными яичниками и с женскими вторичными половыми признаками. Они были способны скрещиваться с нормальными красными самцами. Анализ показал, что красные самки были генотипичеекими самцами. Скрещивание таких самок с нормальными самцами ♀ ХrУR х ♂ ХrУR давало расщепление по полу не 1: 1, а 1 ♀ (XrXr):3 ♂ (2ХrУR и УRУR). Наличие самцов УRУR доказывается результатами скрещивания (см. рис. 123). При действии мужского гормона (метилтестостерона) белые рыбки, т. е. рыбки с генотипом самки - ХrХr становятся самцами. При скрещивании их с нормальными самками (ХrХr) в потомстве получаются только самки.
Рисунок 7. Гормональное переопределение пола у рыб. Гены R - красной, г - белой окраски.
Определение пола у рыб. Определение пола у животных, в том числе у рыб, зависит от наличия половых генов, расположенных либо в половых хромосомах, либо в аутосомах, как это наблюдается у низших позвоночных. Рыбы, с точки зрения формирования пола, представляют разнообразную группу. У одних рыб можно найти кариотипы с половыми хромосомами Х и У, у других - только с Х-хромосомами, а у третьих - половые хромосомы вовсе отсутствуют. Пол у рыб определяется при помощи «главных генов», которые расположены в половых хромосомах. Помимо этого «главные гены», определяющие пол, могут находиться в аутосомах. Половые гены многочисленны, называются они Т-гены, и действуют по принципу полигенов. Различают несколько положений главных генов пола:
У рыб в процессе дифференцировки пола под влиянием факторов внешней и внутренней среды, может произойти переопределение детерминированного генотипом пола. То есть набор половых хромосом не будет соответствовать полу по фенотипу. Эти случаи являются примером функциональных фенотипических инверсий, а особи по полу называются «исключительными». В простом определении - это самки по генотипу, фенотипически являются самцами, и наоборот. Цитогенетические механизмы определения пола у рыб У небольшого числа рыб цитологические методы позволяют выявить половые хромосомы. У таких рыбможно определить каким образом по половым хромосомам генетически детерминируется пол. Так у ерша и окуня обнаружена система определения пола по одной хромосоме ХО, а у угря и моноптеруса гетерогаметный организм уже имеет ХУ, которые можно называть гетерохромосомами. У большинства видов рыб различить половые хромосомы от аутосом трудно, даже если генетический анализ и указывает на наличие гетерохромосом. Вопрос идентификации половых хромосом исследовался рядом авторов, однако они не пришли к единому заключению, потому, что даже у одного вида одни авторы смогли выделить половые хромосомы, другие не подтвердили этого. По всей видимости, отличие половых хромосом от аутосом у рыб незначительное, что и не позволяет многим авторам выделить половые хромосомы цитологическими методами. Генетическая структура хромосом выявляется лучше, чем цитогенетическая. Так в фенотипе у аквариумных рыб мидаки и гуппи очень чётко обнаруживаются гены сцепленные с полом по У-хромосоме. У мидаки существует фактор красной окраски, который передаётся от отца к сыну, локализованный в У-хромосоме, его же аллельный ген, определяющий белую окраску, находится в Х-хромосоме. У гуппи найдено 9 генов сцепленных с У-хромосомой, определяющих окраску и форму плавников. У пецилии также выявлено 5 генов сцепленных с У-хромосомой. Наличие гетерохромосомного механизма определения пола у рыб установлено пока у 4З видов, в том числе у видов с чётко выявляемыми половыми хромосомами, у ерша, угря и окуня, а также у видов, где для выявления половых хромосом применимы генетические методы, в частности анализирующее скрещивание. К таким видам относятся медаки, гуппи, 4 мексиканские расы пецилии и одна раса из Британского Гондураса. Согласно балансной теории пол опредёляется соотношением числа половых хромосом к набору аутосом. Гены, определяющие пол, находятся также и в аутосомах, но они взаимно уравновешивают друг друга и только неравновесные половые гены гетерохромосом отвечают за пол. У рыб балансная теория развита дальше и насколько пересмотрена, так как наблюдается много случаев, когда определение пола происходит под влиянием генов, расположенных в аутосомах. Следовательно, при кроссинговере гены, определяющие пол одного порядка, могут оказаться в одной хромосоме и перекрыть действие генов половых хромосом. Пол таких «исключительных особей» оказывается противоположным определяющему его набору половых хромосом.
Дифференцировка пола у рыб
Половые гены предопределяют развитие половых желез или гонад, которые в свою очередь создают определенный баланс половых гормонов. И фенотипически вторичные половые признаки проявляются за счёт действия гормонов. Если соотношение складывается в пользу андрогенной, развиваются самцы, если в соотношении половых гормонов преобладают эстрогены - получаются самки. Таким образом, каждая клетка тела потенциально бисексуальна иможет даже в течение одной жизни индивидуума, в зависимости от преобладания того или иного полового генома, участвовать в построении признаков характерных то для одного, то для другого пола. У рыб, которые среди позвоночных животных стоят ещё относительно низко на эволюционной лестнице, наблюдается у некоторых видов естественные случаи гермафродитизма. Причем у Serra scriba относящихся к семейству каменных окуней и живущих у нас в Чёрном море, наблюдается функциональный гермафродитизм, когда мужская и женская половые железы развиваются одновременно. Мало того, у них даже наблюдается самооплодотворение. К таким же гермафродитным рыбам относится саргус (рис. 8).
Рисунок 8. Гермафродитные костистые рыбы: вверху - САРГУС; внизу - СОРРАНУС.
Второй вид гермафродитизма у рыб - последовательный гермафродитизм, когда в отдельные периоды жизни особь выступает то как самец, то как самка. Подобный тип гермафродитизма можно встретить у некоторых видов бентосноядных рыб семейства губанов. У истинно раздельнополых рыб, таких как гуппи, можно встретить рудиментарный гермафродитизм. По характеру дифференцировки пола костистых рыб можно разделить на две группы. К первой группе - относятся виды, у которых нейтральная гонада дифференцируется непосредственно в сторону самца или самки. В этой группе наибольшее значение имеют гены, определяющие пол, расположенные непосредственно в половых хромосомах. К таким видам можно отнести медаку, пецилию и других. Ко второй группе - относятся виды, у которых дифференцировка гонады в сторону самца или самки осуществляется опосредованно через первичную гермафродитную ювинальную гонаду. Так определяется и дифференцируется пол у гуппи, меченосца, угря, радужной форели, карпа, макропода. Наличие первичной гермафродитной половой железы может приводить к спонтанным инверсиям пола, когда вместо ожидаемого пола возникает противоположный. Это отмечалось у меченосца, макропода, радужной форели и у других видов. До настоящего времени исследователи ещё не могут установить тот индуктор (наводящее вещество), который недифференцированную гермафродитную гонаду направляют к развитию по пути яичника или семенника. Как уже отмечалось, у высших позвоночных животных гонада несёт корковый или кортикальний и медуллярный (срединный) отделы. Если имеются у животного гонаде эти два слоя, то индукторы кортикальной части гонады заставляют дифференцироваться клетки в яичник, медуллярный же слой держит индукторы заставляющие дифференцироваться клетки в самца. У рыб все осложняется тем, что их гонада не разделена на кортикальную и медиальную части и, следовательно, дифференцировка пола имеет другой механизм. Экспериментальные изучения дифференцировки и инверсии полов с помощью стероидных гормонов, позволили японскому ученому Ямамото высказать мысль, что и в естественных условиях индукторами дифференцировки пола могут служить стероидные половые гормоны. Однако, это предположение ещё гипотетично, так как существуют факторы, определяющие дифференцировку пола у рыб другими воздействиями и даже нейрогенным путём. У рыб дифференцировка гонад контролируется генетическим аппаратом и осуществляется через эндокринную систему зародыша. Однако, на процесс дифференцировки оказывают большое влияние факторы внутренней и внешней среды, которые могут изменить генетически предопределённый пол зародыша. Все это указывает, что механизм определения пола у рыб менее совершенен, чем у других позвоночных животных, нередко можно наблюдать переопределение пола, отсутствует баланс между аутосомами и половыми хромосомами. У-хромосома у рыб, в отличие от высшихпозвоночных, генетически не обеднена, в ней находятся гены ответственные за окраску и ряд других генов сцепленных с полом. Вспомним, что у человека только незначительное число признаков сцепленных с У-хромосомой и передается от отца к сыну. Такие признаки называются голандрическими, например, волосатость мочек ушей, перепонка между пальцами ног. Все же остальные признаки, сцепленные с полом, в том числе и наследственные болезни, передаются через Х-хромосому. У рыб же, У-хромосома, мало отличима от Х-хромосом и несет значительное число генов сцепленных с полом. Малые различия между Х-хромосомой и У-хромосомой у рыб приводят к тому, что между этими хромосомами возможен кроссинговер и обмен локусами хромосом с огромным числом генов. Гены, перешедшие из У-хромосомы в Х-хромосому, наследуются как сцепленные с полом. Резюмируя принципы определения пола у рыб, можно отметить, что в ходе онтогенеза развитие пола определяется генотипом, через воздействие веществ, которые относятся к половым индукторам. Природа половых индукторов пока не установлена. Партеногенез и гиногенез В тесной связи с оогенезом, оплодотворением и определением пола находятся такие явления как партеногенез и гиногенез. Партеногенез (девственное развитие) известен у многих видов животных. У некоторых беспозвоночных, он протекает с гетерогонией, то есть в определённые периоды особи развиваются без самцов и популяция их не имеет, затем появляются из яиц самки и самцы, и наступает период размножения — когда оплодотворение заключается в слиянии сперматозоида и яйцеклетки и, в конечном итоге, в слиянии гаплоидных мужского и женского пронуклеусов, с получением диплоидной зиготы. Гетерогония отмечается, например, у коловраток, тлей и у некоторых других животных (низшие раки). У позвоночных животных экспериментально партеногенез можно получить во всех классах, в том числе и у млекопитающих. В естественных же условиях он обнаружен пока у рыб, амфибий и пресмыкающихся. Так на берегу озера Севан в Армении И.С. Даревский открыл партеногенетический вид ящериц, популяции которых содержат только самок. Созревшая яйцеклетка готова, к оплодотворению, но реакция, сходная с оплодотворением, может произойти либо при каком-нибудь внешнем стимуле, либо под влиянием внутренних факторов, если мы имеем дело с естественным партеногенезом. Все дальнейшее сводится к тому, что без участия сперматозоида происходит кортикальная реакция, возникает оболочка оплодотворения. Если есть кортикальные гранулы, то они буквально выстреливают в оболочку оплодотворения. В процессе кортикальной реакции одно из редукционных телец с гаплоидным набором хромосом втягивается обратно в яйцеклетку, восстанавливается диплоидный набор хромосом, как при оплодотворении. Генотип получившейся особи ничем не отличается от материнского организма, так как несёт с ним одинаковый набор хромосом. Существуют виды партеногенеза без слияния с редукционным тельцем, после кортикальной реакции может начаться развитие гаплоидного организма, который однако, в процессе первых делений восстанавливает диплоидный набор, скорее всего, за счет эндомитоза, когда набор хромосом удваивается, но не образуется веретена деления и удвоившиеся хромосомы не расходятся. Одной из вариаций партеногенеза может считаться гиногенез, хотя некоторые исследователи считают его самостоятельным способом воспроизводства однополых особей. Есть мнение некоторых учёных (Браше, Нарбелъ-Хофстеттер), которые рассматривают гиногенез как одну из переходных стадий к партеногенезу. Гиногенез - развитие только за счет генетического материала, содержащегося в ядре яйцеклетки. При гиногенезе необходимо осеменение сперматозоидами близких видов. Сперматозоиды стимулируют ее к развитию, но слияни
|
||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 1104; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 13.59.82.60 (0.014 с.) |