Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
И биопродуктивность агроэкосистемСодержание книги
Поиск на нашем сайте
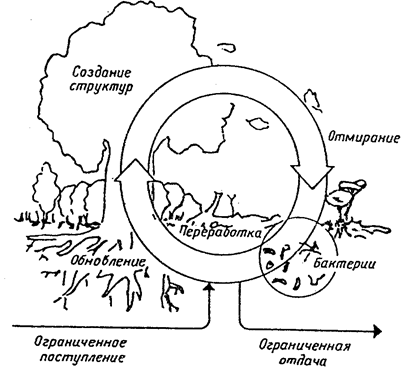
Ранее нами было рассмотрено (глава 4.1), что каждую минуту на 1 см2 верхнего слоя земной атмосферы поступает 2 калории солнечной энергии — так называемая солнечная постоянная, или константа. Использование растениями световой энергии относительно невелико. Только небольшая часть солнечного спектра, так называемая ФАР (фотосинтетически активная радиация с длиной волны 380—710 нм, 21—46% солнечной радиации) используется в процессе фотосинтеза. В зоне умеренного климата на сельскохозяйственных землях КПД фотосинтеза не превышает 1,5—2%, а чаще всего он равен 0,5%. В развивающемся мировом сельском хозяйстве различаются по количеству поступающей и используемой человеком энергии и ее источнику несколько типов экосистем (М.С. Соколов и др 1994). 1. Естественные экосистемы. Единственным источником энергии является солнечная (океан, горные леса). Эти экосистемы представляют собой основную опору жизни на Земле (приток энергии в среднем 0,2 ккал/см2 • год). 2. Высокопродуктивные естественные экосистемы. Кроме солнечной, используются другие естественные источники энергии (каменный уголь, торф и т. д.). К ним относятся лиманы, дельты крупных рек, влажные тропические леса и другие естественные экосистемы, обладающие высокой продуктивностью. Здесь в избытке синтезируется органическое вещество, которое используется или накапливается (приток энергии в среднем 2 ккал/см2 • год). 3. Агроэкосистемы, близкие к естественным экосистемам. Наряду с солнечной энергией используются дополнительные источники, создаваемые человеком. Сюда относятся системы сельского и водного хозяйства, которые производят продовольствие и сырье. Дополнительные источники энергии — ископаемое топливо, энергия обмена веществ людей и животных (приток энергии в среднем 2 ккал/см2 • год). 4. Агроэкосистемы интенсивного типа. Связаны с потреблением больших количеств нефтепродуктов и агрохимикатов. Они более продуктивны в сравнении с предыдущими экосистемами, отличаясь высокой энергоемкостью (приток энергии в среднем 20 ккал/см2 • год). 5. Промышленные (городские) экосистемы. Получают готовую энергию (газ, уголь, электричество). К ним относятся города, пригородные и промышленные зоны. Они являются как генераторами улучшения жизни, так и источниками загрязнения среды (поскольку прямая солнечная энергия не используется): Эти системы биологически связаны с предыдущими. Промышленные экосистемы очень энергоемкости (приток энергии в сред-' нем 200 ккал/см2 • год). Основные отличительные особенности функционирования природных экосистем и агроэкосистем. 1. Разное направление отбора. Для природных экосистем xaрактерен естественный отбор, который ведет к фундаментальному их свойству — устойчивости, отметая неустойчивые, нежизнеспособные формы организмов их сообществ. Агроэкосистемы создаются и поддерживаются человеком. Главным здесь является искусственный отбор, который направлен на повышение урожайности сельскохозяйственных культур. Нередко урожайность сорта не связана с его устойчивостью к факторам окружающей среды, вредными организмами. 2. Разнообразие экологического состава фитоценоза обеспечивает устойчивость продукционного состава в естественной экосистеме при колебании в различные годы погодных условий. Угнетение одних видов растений приводит к повышению продуктивности других. В результате фитоценоз и экосистема в целом сохраняет способность к созданию определенного уровня продукции в разные годы. Агроценоз полевых культур — сообщество монодоминантное, а нередко и односортовое. На всех растениях агроценоза действие неблагоприятных факторов отражается одинаково. Не может быть компенсировано угнетение роста и развития основной культуры усиленным ростом других видов растений. И как результат, устойчивость продуктивности агроценоза ниже, чем в естественных экосистемах. 3. Наличие разнообразия видового состава растений с различными фенологическими ритмами дает возможность фитоценозу как целостной системе осуществлять непрерывно в течение всего вегетационного периода продукционный процесс, полно и экономно расходуя ресурсы тепла, влаги и питательных элементов. Период вегетации культивируемых растений в агроценозах короче вегетационного сезона. В отличие от естественных фитоценозов, где виды различного биологического ритма достигают максимальной биомассы в разное время вегетационного сезона, в агроценозе рост растений одновременен и последовательность стадий развития, как правило, синхронизирована. Отсюда, время взаимодействия фитоком-понента с другими компонентами (например, почвой) в агроценозе намного короче, что, естественно, сказывается на интенсивности обменных процессов в целой системе. Разновременность развития растений в естественной (природной) экосистеме и одновременность их развития в агроценозе приводят к различному ритму продукционного процесса. Ритм продукционного процесса, например, в естественных лугопастбищных экосистемах, задает ритм деструкционным процессам или определяет скорость минерализации растительных остатков и время ее максимальной и минимальной интенсивности. Ритм дест-рукционных процессов в агроценозах в значительно меньшей степени зависит от ритма продукционного процесса, ввиду того что наземные растительные остатки поступают на почву и в почву на короткий промежуток времени, как правило, в конце лета и в начале осени, а их минерализация осуществляется главным образом на следующий год. 4. Существенным различием естественных экосистем и агроэкосистем является степень скомпенсированности круговорота веществ внутри экосистемы. Круговороты веществ (химических элементов) в естественных экосистемах осуществляются по замкнутым циклам или близки к скомпенсированности: приход вещества в цикл за определенный период в среднем равен выходу вещества из цикла, а отсюда внутри цикла приход вещества в каждый блок приблизительно равен вькоду вещества из него (рис. 18.5).
Рис. 18.5. Круговорот питательных веществ в естественной экосистеме (по А. Тарабрину, 1981)
Антропогенные воздействия нарушают замкнутость круговорота веществ в экосистемах (рис. 18.6).
Рис. 18.6. Круговорот питательных веществ в агроэкосистеме (по А. Тарабрину, 1981) Часть вещества в агроценозах безвозвратно изымается из экосистемы. При высоких нормах внесения удобрений для отдельных элементов может наблюдаться явление, когда величина входа элементов питания в растения из почвы оказывается меньше величины поступления элементов питания в почву из разлагающихся растительных остатков и удобрений. С хозяйственно полезной продукцией в агроценозах отчуждается 50—60% органического вещества от его количества, аккумулированного в продукции. 5. Природные экосистемы являются системами, если можно так выразиться, авторегуляторными, а агроценозы — управляемые человеком. Для достижения своей цели человек в агроценозе изменяет или контролирует в значительной мере влияние природных факторов, дает преимущества в росте и развитии, главным образом компонентам, которые продуцируют пищу. Основная задача в связи с этим — найти условия повышения урожайности при минимализации энергетических и вещественных затрат, повышении почвенного плодородия. Решение данной задачи состоит в наиболее полном использовании агрофитоценозами природных ресурсов и создании скомпенсированных циклов химических элементов в агроценозах. Полнота использования ресурсов определяется генетическими особенностями сорта, продолжительностью вегетации, неоднородностью компонентов в совместных посевах, ярусностью посева и т. д. Следовательно, делает вывод М.С. Соколов и др. (1994), самый строгий контроль состояния агроэкосистем, который требует значительных затрат энергии, можно осуществить только в закрытом пространстве. К данной категории относят полуоткрытые системы с весьма ограниченными каналами сообщения с внешней средой (теплицы, животноводческие коплексы), где регулируются и в значительной степени контролируются температура, радиация, круговорот минеральных и органических веществ. Это — управляемые агроэкосис-темы. Все другие агроэкосистемы — открытые. Со стороны человека эффективность контроля тем выше, чем они проще. В полуоткрытые и открытых системах усилия человека сводятся к обеспечению оптимальных условий роста организмов и строгому биологическому контролю за их составом. Исходя из этого возникают следующие практические задачи: — во-первых, по возможности полное устранение нежелательных видов; — во-вторых, отбор генотипов, обладающих высокой потенциальной продуктивностью. В целом круговорот веществ связывает различные виды, населяющие а^оэкосистемы (рис. 18.7).
Рис. 18.7. Поток энергии в пастбищной агроэкосистеме (по Н.А. Уразаеву и др., 1996): Примечание: белыми стрелками показана миграция веществ от продуцентов к первичным и вторичным консументам, черными — минерализация органических остатков растений и животных
Автотрофные организмы — продуценты, главным образом травы (I); первичные консументы, большей частью сельскохозяйственные животные (II); вторичные консументы — паразиты и микроорганизмы (III) и организмами-редуцентами являются грибы и микробы (IV). Отдельные живые организмы (животные) по отношению к звеньям трофической цепи было бы неправильным рассматривать только как консументы, а микроорганизмы как исключительно редуцен-ты и деструкторы. Утилизируя органические соединения, животные разлагают их до простейших соединений — аммиака, мочевины, углекислого газа, воды или выступают как редуценты. Микроорганизмы, поедаемые хищньми простейшими, выступают как пищевой субстрат и источник энергии для консументов и т. д. В биосфере многие циркулирующие вещества биогенного происхождения одновременно являются и носителями энергии. Растения в процессе фотосинтеза превращают лучистую энергию Солнца в энергию химических связей органических веществ и накапливают ее в форме углеводов — потенциальных энергоносителей. Данная энергия включается в круговорот питания от растений через фитофаги к консументам более высоких порядков. Количество связанной энергии по мере движения по трофической цепи постоянно уменьшается, так как значительная ее часть расходуется для поддержания жизненных функций консументов. Благодаря круговороту энергии в экосистеме поддерживается разнообразие форм жизни, а система сохраняет устойчивость. По М.С. Соколову и др. (1994) расход фотосинтетической энергии растений в агроэкосистеме на примере лугопастбищных угодий средней полосы России выглядит следующим образом: — около 1/6 части используемой растениями энергии расходуется на дыхание; — около 1/4 части энергии поступает в организм растительнояд-ных животных. При этом 50% ее оказывается в экскрементах и трупах животных; — в целом вместе с отмершими растениями и фитофагами около 3/4 первоначально поглощенной энергии содержится в мертвом органическом веществе и немногим более 1/4 исключается из экосистемы при дыхании в форме тепла. Еще раз отметим, что поток энергии в пищевой цепи агроэко-системы подчиняется закону превращения энергии в экосистемах, так называемому закону Линдемана, или закону 10%. По закону Линдемана, только часть энергии, поступившей на определенный трофический уровень агроценоза (биоценоза), передается организмам, находящимся на более высоких трофических уровнях (рис. 18.8).
Рис. 18.8. Потери энергии в пищевой цепи (по Т. Миллеру, 1994)
Передача энергии с одного уровня на другой происходит с очень малым КПД. Этим объясняется ограниченное количество звеньев в пищевой цепи независимо от того или иного агроценоза. Количество энергии, продуцируемое в конкретной природной экосистеме, является довольно стабильной величиной. Благодаря способности экосистемы производить биомассу, человек получает необходимые ему пищевые и многие технические ресурсы. Как уже было отмечено, проблема обеспечения численно растущего человечества пищей — это главным образом проблема повышения продуктивности агроэкосистем (сельского хозяйства), рис. 18.9.
Рис.18.9. Блок-схема продуктивности агроэкосистем Воздействие человека на экологические системы, связанное с их разрушением или загрязнением, непосредственно ведет к прерыванию потока энергии и вещества, а значит, и к снижению продуктивности. Поэтому первая задача, стоящая перед человечеством, — предотвращение снижения продуктивности агроэкосистем, а после ее решения может быть решена и вторая важнейшая задача — повышение продуктивности. В 90-х гг. XX в. годовая первичная продуктивность обрабатываемых земель на планете составляла 8,7 млрд т, а запас энергии — 14,7×1017кДж. Отношения организмов в Агроэкосистемах
Составными частями агроэкосистемы являются сельскохозяйственные угодья, на которых выращиваются зерновые, пропашные, кормовые и технические культуры, а также луга и пастбища. Основными элементами агробиоценоза в аграрных экосистемах являются (по М. В. Маркову, 1972): 1. Культурные растения, высеянные или высаженные человеком. 2. Сорные растения, которые проникли в агробиоценоз помимо, а иногда и вопреки воле человека. 3. Микроорганизмы ризосфер культурных и сорных растений. 4. Клубеньковые бактерии на корнях бобовых, связывающие свободный азот воздуха. 5. Микоризообразующие грибы на корнях высших растений. 6. Бактерии, грибы, актиномицеты, водоросли, свободно живущие в почве. 7. Беспозвоночные животные, живущие в почве и на растениях. 8. Позвоночные животные (грызуны, птицы и др.), живущие в почве и посевах. 9. Грибы, бактерии, вирусы — паразиты (полупаразиты) культурных и сорных растений. 10. Бактериофаги — паразиты микроорганизмов. Агроэкосистема обладает биологической продуктивностью или биологической емкостью. Размер популяций отдельных входящих в них видов колеблется из-за постоянных изменений абиотических и биотических факторов. К факторам, оказывающим влияние на плотность популяции вида, относится межвидовая конкуренция в отношении пищи и пространства. Межвидовая конкуренция возникает главным образом тогда, когда у разных видов имеются одинаковые или близкие к условиям среды требования. При увеличивающемся недостатке средств существования конкуренция усиливается. Обычно плотность популяций различных групп организмов в агроэкосистеме поддерживается на оптимальном уровне. В агрофитоценозе регулирование плотности популяций прояв-, ляется в виде внутривидовой конкуренции растений, и как результат, устанавливается их относительная оптимальная плотность на занятой территории. Например, число растений клевера на 1 м2 к моменту уборки покровной культуры составляет 400 шт./м2. В следующем году к началу вегетации оно может снизиться до 150—200 шт./м2, что создает наиболее благоприятные условия для формирования урожая. Регуляция плотности растительного покрова также происходит под влиянием таких факторов, как плотность листовой поверхности, выраженная через индекс ассимилирующей поверхности. Обостряется конкуренция при высокой плотности листовой поверхности. Так как не все растения получают достаточное количество света, более слабые подавляются. Следовательно, между особями одного и того же вида наблюдается внутривидовая конкуренция. Величина популяции вида ограничивается величиной необходимых для ее жизни ресурсов окружающей среды. Межвидовая конкуренция растений не приводит к полному вытеснению менее конкурентоспособного вида. Как процесс борьбы между культурными и сорными растениями, проявляется межвидовая конкуренция в открытой агроэкосистеме. На лугах и пастбищах такая форма конкуренции преобладает. Растительные сообщества здесь характеризуются типичными особенностями, свойственными данной территории. Посевы культурных растений в агрофитоценозе являются единственным источником питания для травоядных животных и насекомых-фитофагов. В благоприятные для роста растений периоды популяции продуцентов могут резко и быстро увеличиваться. Обычно наносит большой ущерб сельскохозяйственным культурам массовое размножение травоядных и насекомых- фитофагов. Естественное регулирование численности травоядных животных, насекомых-фитофагов и доведение их популяций до экономически безвредного порога путем использования их естественных врагов-хищников сложно и не всегда дает хорошие результаты. Отсюда в сельскохозяйственной практике искусственное вмешательство и регулирование численности фитофагов осуществляется за счет использования различных искусственных средств защиты. Анализ основных трофических цепей в агроэкосистеме обычно показывает, что биофаги (фитофаги, хищники, паразиты) активно влияют на собственную численность путем частичного использования или разрушения предшествующего звена трофической цепи, которое служит им источником энергии. Биофаги путем преобразования поглощенных веществ создают специфические источники энергии для последующих звеньев: ткани собственного тела — для биофагов, экскременты — для капрофагов, трупы — для некрофагов. Таким образом, биофаги (сапрофаги) пассивно определяют энергетический обмен у сменяющих их консументов. Совместная и многосторонняя деятельность самых разных организмов экосистемы, в первую очередь гетеротрофов, препятствует длительному накоплению мертвого органического вещества с заключенной в нем химической энергией. Под влиянием фитофагов снижение продуктивности растений не всегда пропорционально количеству потребляемой ими пищи, их доминированию или биомассе, а обусловлено характером повреждения автотрофов, их возрастом и состоянием. Например, если фитофаг нападает на молодое растение, то в некоторых случаях наносится больший ущерб, чем при питании на взрослых растениях (крестоцветные блошки и др.). Напротив, в других случаях молодые растения успешнее способны компенсировать ущерб за счет образования новых побегов или более интенсивного роста здоровых побегов, чем растения, пострадавшие в более поздние сроки. Нередко ущерб, причиненный животными, уравновешивается приносимой ими пользой. Так, грачи при выкармливании потомства уничтожают вредителей сельскохозяйственных культур, и в то же время могут наносить ущерб, повреждая всходы кукурузы, зерновых культур. В целом же следует еще раз отметить, что в агроэкосистемах пищевые цепи вовлечены в сферу деятельности человека. В них изменена экологическая пирамида. На вершине экологической пирамиды стал человек (рис. 18.10).
Рис. 18.10. Пищевая пирамида (по П. Фарбу, 1971) Пояснения: при переходе от одного трофического уровня к другому происходит потеря 9/10 живого веса. Так, из 400 кг растительного планктона можно получить 40 кг животного планктона и соответственно 4 кг рыбы — количество, необходимое человеку, чтобы прибавить в весе на 0,4 кг.
Своеобразие экологической пирамиды, на вершине которой находится человек, — специфический признак любой агроэкосистемы. В агро-экосистемах видовой состав растений и животных обеднен. Аграрные экосистемы малокомпонентны. Малокомпонентность также один из признаков агроэкосистемы. Ландшафтная организация Агроэкосистем
В конце XX в. наиболее широкое распространение получило следующее определение ландшафта. Ландшафт — это имеющий естественные границы участок зеленой поверхности, в пределах которого все природные компоненты (горные породы, рельеф, воды, почвы, растительный и животный мир) образуют взаимосвязанное и исторически взаимообусловленное единство. При этом каждый ландшафт характеризуется также определенным уровнем прихода разных видов энергии и параметрами атмосферно-климатических процессов, показателями и динамикой круговорота веществ. Классификация ландшафтов по А.А. Иноземцову, Ю.А. Щербакову (1988) построена на основе зональных и азональных признаков в дифференции природы поверхности суши (рис. 18.11).
Рис. 18.11. Соотношение таксономических единиц физико-географического районирования суши (по А. А. Иноземцеву, Ю. А. Щербакову, 1988)
Полная система классификации ландшафтов, основанная на учете, как зональных, так и азональных признаков, строится по принципу: страна ® зона ® провинция ® подзона и т. д., т. е. путем чередования единиц, выделенных на основании зональных и азональных признаков. Основные показатели потенциала возобновимых ресурсов по некоторым типам равнинных ландшафтов европейской части России представлены в табл. 18.1. В основе формирования потенциала возобновимых ресурсов лежит соотношение обеспеченности ландшафта теплом и влагой. По мере продвижения с севера на юг происходит нарастание тепловых ресурсов, увеличивается и биопродуктивность ландшафтов. Таблица 18.1
|
||
|
|
Последнее изменение этой страницы: 2016-04-20; просмотров: 531; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.108 (0.01 с.) |