Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Индекс интенсивности биологического круговоротаСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
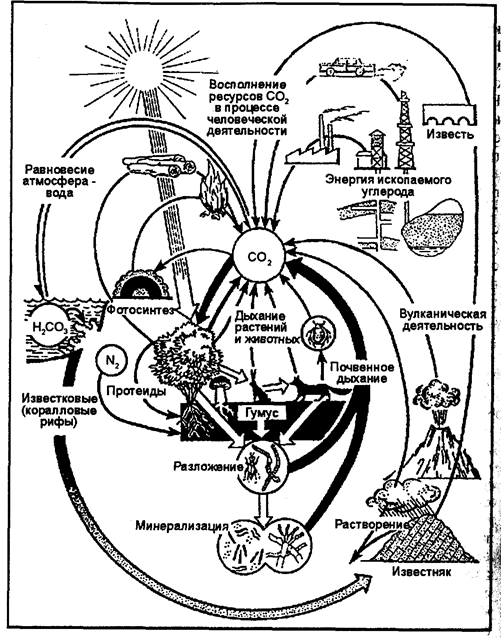
Круговорот углерода. Из всех биогеохимических циклов круговорот углерода, без сомнения, самый интенсивный. С высокой скоростью углерод циркулирует между различными неорганическими средствами и через посредство пищевых сетей внутри сообществ живых организмов (рис. 12.14).
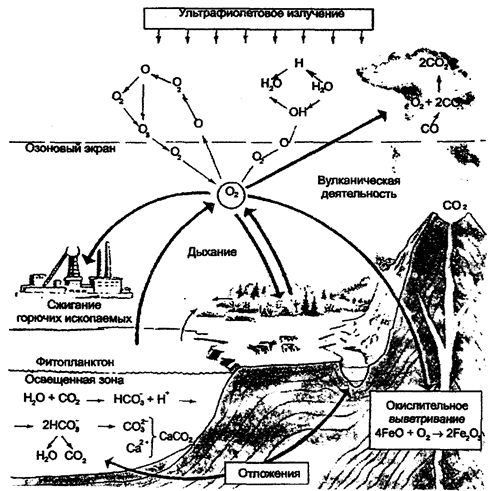
Рис. 12.14. Круговорот углерода (по И. П. Герасимову, 1980) В круговороте углерода определенную роль играют СО и СО2. Часто в биосфере Земли углерод представлен наиболее подвижной формой СО2. Источником первичной углекислоты биосферы является вулканическая деятельность, связанная вековой дегазацией мантии и нижних горизонтов земной коры. Миграция СО2 в биосфере протекает двумя путями. Первый путь заключается в поглощении его в процессе фотосинтеза с образованием глюкозы и других органических веществ, из которых построены все растительные ткани. В дальнейшем они переносятся по пищевым цепям и образуют ткани всех остальных живых существ экосистемы. Следует заметить, что вероятность отдельно взятого углерода «побывать» в течение одного цикла в составе многих организмов мала, потому что при каждом переходе с одного трофического уровня на другой велика возможность, что содержащая его органическая молекула будет расщеплена в процессе клеточного дыхания для получения энергии. Атомы углерода при этом вновь поступают в окружающую среду в составе углекислого газа, таким образом завершив один цикл и приготовившись начать следующий. В пределах суши, где имеется растительность, углекислый газ атмосферы в процессе фотосинтеза поглощается в дневное время. В ночное время часть его выделяется растениями во внешнюю среду. С гибелью растений и животных на поверхности происходит окисление органических веществ с образованием СО2. Атомы углерода возвращаются в атмосферу и при сжигании органического вещества. Важная и интересная особенность круговорота углерода состоит в том, что в далекие геологические эпохи, сотни миллионов лет назад, значительная часть органического вещества, созданного в процессах фотосинтеза, не использовалась ни консументами, ни редуцентами, а накапливалась в литосфере в виде ископаемого топлива; нефти, угля, горючих сланцев, торфа и др. Это ископаемое топливо добывается в огромных количествах для обеспечения энергетических потребностей нашего индустриального общества. Сжигая его, мы в определенном смысле завершаем круговорот углерода. По второму пути миграция углерода осуществляется созданием карбонатной системы в различных водоемах, где СО2 переходит в Н2СО3, НСО13, СО23. С помощью растворенного в воде кальция (или магния) происходит осаждение карбонатов (СаСО3) биогенным и абиогенным путями. Образуются мощные толщи известняков. По А. Б. Ронову, отношение захороненного углерода в продуктах фотосинтеза к углероду в карбонатных породах составляет 1:4. Существует наряду с большим круговоротом углерода и ряд малых его круговоротов на поверхности суши и в океане. В целом же без антропогенного вмешательства содержание углерода в биогеохимических резервуарах: биосфере (биомасса+почва и детрит), осадочных породах, атмосфере и гидросфере, — сохраняется с высокой степенью постоянства. По Т.А. Акимовой, В.В. Хаскину (1994), постоянный обмен углеродом, с одной стороны, между биосферой, а с другой — между атмосферой и гидросферой, обусловлен газовой функцией живого вещества — процессами фотосинтеза, дыхания и деструкции, и составляет около 6×1010 т/год. Существует поступление углерода в атмосферу и гидросферу и при вулканической деятельности в среднем 4,5×106 т/год. Общая масса углерода в ископаемом топливе (нефть, газ, уголь и др.) оценивается в 3,2×1015 т, что соответствует средней скорости накопления 7 млн т/год. Это количество по сравнению с массой циркулирующего углерода незначительное и как бы выпадало из круговорота, терялось в нем. Отсюда степень разомкну-гости (несовершенства) круговорота составляет 10-4, или 0,01%, а соответственно степень замкнутости — 99,99%. Это означает, с одной стороны, что каждый атом углерода принимал участие в цикле десятки тысяч раз, прежде чем выпал из круговорота, оказался в недрах. А с другой стороны — потоки синтеза и распада органических веществ в биосфере с очень высокой точностью подогнаны друг к другу. В. Г. Горшковым (1988) на основе расчетов делается важное заключение: «Потоки синтеза и разложение органических веществ совпадают с точностью 10"4 и скоррелированы с точностью 10-4. Скоррелированность потоков синтеза и распада с указанной точностью доказывает наличие биологической регуляции окружающей среды, ибо случайная связь величин с такой точностью в течение миллионов лет невероятна». В постоянном круговороте находится 0,2% мобильного запаса углерода. Углерод биомассы обновляется за 12, атмосферы — за восемь лет. Огромный контраст между краткостью данных периодов, постоянством и возрастом биосферы подтверждает высочайшую сбалансированность «мира углерода». Круговорот кислорода. Кислород (О2) играет важную роль в жизни большинства живых организмов на нашей планете. В количественном отношении это главная составляющая живой материи. 349 Например, если учитывать воду, которая содержится в тканях, то тело человека содержит 62,8% кислорода и 19,4% углерода. В целом в биосфере этот элемент по сравнению с углеродом и водородом является основным среди простых веществ. В пределах биосферы происходит быстрый обмен кислорода с живыми организмами или их остатками после гибели. Растения, как правило, производят свободный кислород, а животные являются его потребителями путем дыхания. Будучи самым распространенным и подвижным элементом на Земле, кислород не лимитирует существование и функции экосферы, хотя доступность кислорода для водных организмов может временно и ограничиться. Круговорот кислорода в биосфере необычайно сложен, так как с ним в реакцию вступает большое количество органических и неорганических веществ. В результате возникает множество эпициклов, происходящих между литосферой и атмосферой или между гидросферой и двумя этими средами. Круговорот кислорода в некотором отношении напоминает обратный круговорот углекислого газа. Движение одного происходит в направлении, противоположном движению другого (рис. 12.15). Потребление атмосферного кислорода и его возмещение первичными продуцентами происходит сравнительно быстро. Так, для полного обновления всего атмосферного кислорода требуется 2000 лет. В наше время фотосинтез и дыхание в природных условиях, без учета деятельности человека, с большой точностью уравновешивают друг друга. В связи с этим накопления кислорода в атмосфере не происходит, и его содержание (20,946%) остается постоянным.
Рис. 12.15. Круговорот кислорода (по Е. А. Криксунову и др., 1995)
В верхних слоях атмосферы при действии ультрафиолетовой радиации на кислород образуется озон — О3: hv ® О2 «2О; О + О «О3; DН = +141,9 кДж/моль. Здесь hv — квант света с длиной волны не более 225 нм.
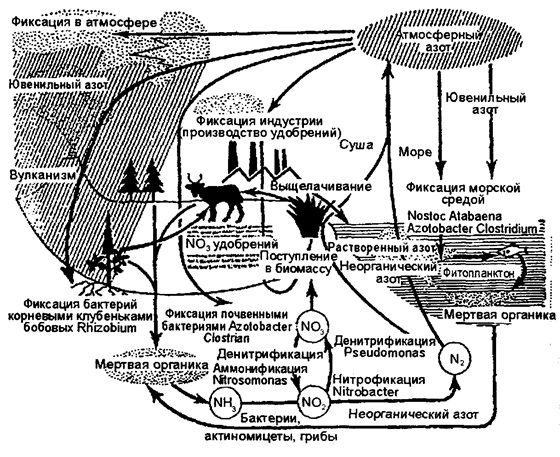
На образование озона тратится около 5% поступающей к Земле солнечной энергии — около 8,6×1015 Вт. Реакции легко обратимы. При распаде озона эта энергия выделяется, за счет чего в верхних слоях атмосферы поддерживается высокая температура. Средняя концентрация озона в атмосфере составляет около 10-6 об. %; максимальная концентрация О3 —до 4×10-6 об. % достигается на высотах 20—25 км (ТА. Акимова, В.В. Хаскин (1998). Озон служит своеобразным УФ-фильтром: задерживает значительную часть жестких ультрафиолетовых лучей. Вероятно, образование озонового слоя было одним из условий выхода жизни из океана и заселения суши. Большая часть кислорода, вырабатываемого в течение геологических эпох, не оставалась в атмосфере, а фиксировалась литосферой в виде карбонатов, сульфатов, окислов железа и т. п. Эта масса составляет 590×1014 т пpoтив39×1014 т киcлopoдa, который циркулирует в биосфере в виде газа или сульфатов, растворенных в континентальных и океанических водах. Круговорот азота. Азот — незаменимый биогенный элемент, так как он входит в состав белков и нуклеиновых кислот. Круговорот азота один из самых сложных, поскольку включает как газовую, так и минеральную фазу, и одновременно самых идеальных круговоротов (рис. 12.16).
Рис. 12.16. Круговорот азота (по Ф. Рамаду, 1981)
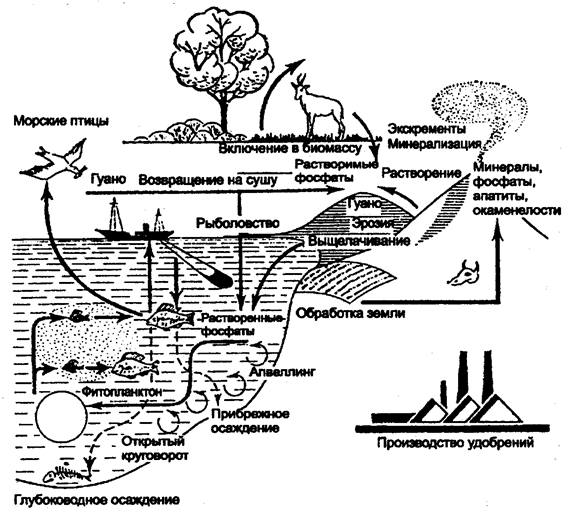
Круговорот азота тесно связан с круговоротом углерода. Как правило, азот следует за углеродом, вместе с которым он участвует в образовании всех протеиновых веществ. Атмосферный воздух, содержащий 78% азота, является неисчерпаемым резервуаром. Однако основная часть живых организмов не может непосредственно использовать этот азот. Он должен быть предварительно связан в виде химических соединений. Например, для усвоения азота растениями необходимо, чтобы он входил в состав ионов аммония (NH4+) или нитрата (NO3-). Газообразный азот непрерывно поступает в атмосферу в результате работы денитрофицирующих бактерий, а бактерии-фиксаторы вместе с сине-зелеными водорослями (цианофитами) постоянно поглощают его, преобразуя в нитраты. Важную роль в превращении газообразного азота в аммонийную форму в ходе так называемой азотофиксации играют бактерии из рода Rhizobium, живущие в клубеньках на корнях бобовых растений. Растения обеспечивают бактерий местообитанием и пищей (сахара), получая взамен от них доступную форму азота. По пищевым цепям органический (входящий в состав органических молекул) азот передается от бобовых другим организмам экосистемы. В процессе клеточного дыхания белки и другие содержащие азот органические соединения расщепляются, азот выделяется в среду большей частью в аммонийной форме (NH4+). Некоторые бактерии способны переводить ее и в нитратную (NO3-) форму. Отметим, что обе эти формы азота усваиваются любыми растениями. Азот, таким образом, совершает круговорот как минеральный биоген. Однако такая минерализация обратима, так как почвенные бактерии постоянно превращают нитраты снова в газообразный азот. В водной среде также существуют различные виды нитрофи-цирующих бактерий, но главная роль в фиксации атмосферного азота здесь принадлежит многочисленным видам способных к фотосинтезу сине-зеленых водорослей из родов Anabaena, Nostoc, Frichodesmium и др. Круговорот азота четко прослеживается и на уровне деструкторов. Протеины и другие формы органического азота, содержащиеся в растениях и животных после их гибели, подвергаются воздействию гетеротрофных бактерий, актиномицетов, грибов (биоредуцирующих микроорганизмов), которые вырабатывают необходимую им энергию восстановлением этого органического азота, преобразуя его таким образом в аммиак. В почвах происходит процесс нитрификации, состоящий из цепи реакций, где при участии микроорганизмов осуществляется окисление иона аммония (МН4+) до нитрита (NO2-) или нитрита до нитрата (NО3-). Восстановление нитритов и нитратов до газообразных соединений молекулярного азота (N2) или окиси азота (N2O) составляет сущность процесса денитрификации. Образование нитратов неорганическим путем в небольших количествах постоянно происходит и в атмосфере: путем связывания атмосферного азота с кислородом в процессе электрических разрядов во время гроз, а затем выпадением с дождями на поверхность почвы. Еще одним источником атмосферного азота являются вулканы, компенсирующие потери азота, выключенного из круговорота при седиментации или осаждении его на дно океанов. В целом же среднее поступление нитратного азота абиотического происхождения при осаждении из атмосферы в почву не превышает 10 кг (год/га), свободные бактерии дают 25 кг (год/га), в то время как симбиоз Rhizobium с бобовыми растениями в среднем продуцирует 200 кг (год/га). Преобладающая часть связанного азота перерабатывается денитрифицирующими бактериями в N и вновь возвращается в атмосферу. Лишь около 10% аммонифицированного и нитрифицированного азота поглощается из почвы высшими растениями и оказывается в распоряжении многоклеточных представителей биоценозов. Круговорот фосфора. Круговорот фосфора в биосфере связан с процессами обмена веществ в растениях и животных. Этот важный и необходимый элемент протоплазмы, содержащийся в наземных растениях и водорослях 0,01—0,1%, животных от 0,1% до нескольких процентов, циркулирует, постепенно переходя из органических соединений в фосфаты, которые снова могут использоваться растениями (рис. 12.17).
Рис. 12.17. Круговорот фосфора (по Ф. Рамаду, 1981)
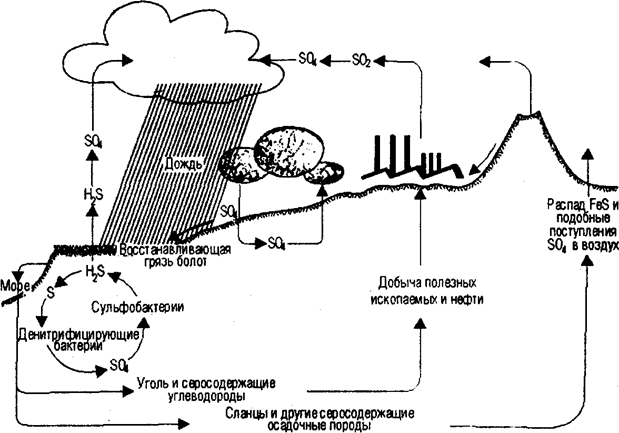
Однако фосфор в отличие от других биофильных элементов в процессе миграции не образует газовой формы. Резервуаром фосфора является не атмосфера, как у азота, а минеральная часть литосферы. Основными источниками неорганического фосфора являются изверженные породы (апатиты) или осадочные породы (фосфориты). Из пород неорганический фосфор вовлекается в циркуляцию выщелачиванием и растворением в континентальных водах. Попадая в экосистемы суши, почву, фосфор поглощается растениями из водного раствора в виде неорганического фосфат-иона (РО43-) и включается в состав различных органических соединений, где он выступает в форме органического фосфата. По пищевым цепям фосфор переходит от растений к другим организмам экосистемы. Химически связанный фосфор попадает с остатками растений и животных в почву, где вновь подвергается воздействию микроорганизмов и превращается в минеральные ортофосфаты, а в дальнейшем происходит повторение цикла. В водные экосистемы фосфор переносится текучими водами. Реки непрерывно обогащают фосфатами океаны. В соленых морских водах фосфор переходит в состав фитопланктона, служащего пищей другим организмам моря, в последующем накапливаясь в тканях морских животных, например рыб. Часть соединений фосфора мигрирует в пределах небольших глубин, потребляясь организмами, другая часть теряется на больших глубинах. Отмершие остатки организмов приводят к накоплению фосфора на разных глубинах. Отсюда следует, что фосфор, попадая в водоемы тем или иным путем, насыщает, а нередко и перенасыщает их экосистемы. Частичный возврат фосфатов на сушу связан с поднятием земной коры выше уровня моря. Определенное количество фосфора переносится на сушу морскими птицами, а также благодаря рыболовству. Птицы отлагают фосфор на отдельных островах в виде гуано. При рассмотрении круговорота фосфора в масштабе биосферы за сравнительно короткий период можно отметить, что он полностью не замкнут. Механизм возвращения фосфора из океанов на сушу в естественных условиях совершенно не способен компенсировать потери этого элемента на седиментацию. Круговорот серы. Существуют многочисленные газообразные соединения серы, такие, как сероводород H-S и сернистый ангидрид SO2. Однако преобладающая часть круговорота этого элемента имеет осадочную природу и происходит в почве и воде. Основной источник серы, доступный живым организмам, — сульфаты (SO4,). Доступ неорганической серы в экосистеме облегчает хорошая растворимость многих сульфатов в воде. Растения, поглощая сульфаты, восстанавливают их и вырабатывают серосодержащие аминокислоты (метионин, цистеин, цистин), играющие важную роль в выработке третичной структуры протеинов при формировании дисульфидных мостиков между различными зонами полипептидной цепи. Подробная схема круговорота серы приведена на рис. 12.18. Здесь хорошо просматриваются многие основные черты биогеохимического круговорота. 1. Обширный резервный фонд в почве и отложениях, меньший в атмосфере. 2. Ключевую роль в быстро обменивающемся фонде играют специализированные микроорганизмы, выполняющие определенные реакции окисления или восстановления. Благодаря процессам окисления и восстановления происходит обмен серы между доступными сульфатами (SO4) и сульфидами железа, находящимися глубоко в почве и осадках. Специализированные микроорганизмы выполняют реакции: H2S ® S ® SO4 — бесцветные, зеленые и пурпурные серобактерии; SO4 ® H2S (анаэробное восстановление сульфата) — Desulfovibrio; H2S ® SO4(аэробное окисление сульфида) — тиобациллы; органическая S в SO4 и H2S — аэробные и анаэробные гетеротрофные микроорганизмы соответственно. Первичная продукция обеспечивает включение сульфата в органическое вещество, а экскреция животными служит путем возвращения сульфата в круговорот. 3. Микробная регенерация из глубоководных отложений, приводящая к движению вверх газовой фазы H2S.
Рис. 12.18. Круговорот серы (по Ф. Рамаду, 1981)
4. Взаимодействие геохимических и метеорологических процессов — эрозия, осадкообразование, выщелачивание, дождь, абсорбация-десорбция и др. — с такими биологическими процессами, как продукция и разложение. 5. Взаимодействие воздуха, воды и почвы в регуляции кругом ворота в глобальном масштабе. В целом экосистеме по сравнению с азотом и фосфором требуется меньше серы. Отсюда сера реже является лимитирующим фактором для растений и животных. Вместе с тем круговорот серы относится к ключевым в общем процессе продукции и разложения биомассы. К примеру, при образовании в осадках сульфидов железа фосфор из нерастворимой формы переводится в растворимую и становится доступным для организмов. Это подтверждение того, как один круговорот регулируется другим. Поток энергии в экосистемах
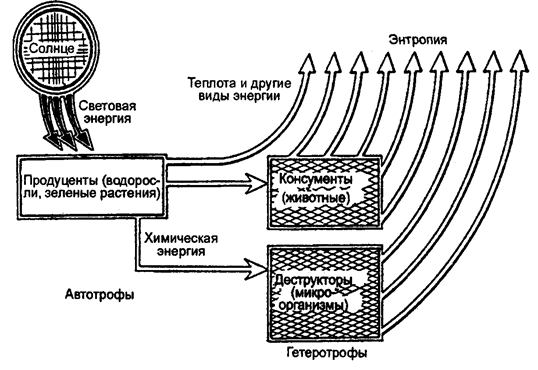
Поддержание жизнедеятельности организмов и круговорот вещества в экосистемах, т. е. существование экосистем, зависит от постоянного притока энергии, необходимой всем организмам для их жизнедеятельности и самовоспроизведения (рис. 12.19).
Рис. 12.19. Поток энергии в экосистеме (по Ф. Рамаду, 1981)
В отличие от веществ, непрерывно циркулирующих по разным блокам экосистемы, которые всегда могут повторно использоваться, входить в круговорот, энергия может быть использована только раз, т. е. имеет место линейный поток энергии через экосистему. Одностороний приток энергии как универсальное явление природы происходит в результате действия законов термодинамики. Первый закон гласит, что энергия может превращаться из одной формы (например, света) в другую (например, потенциальную энергию пищи), но не может быть создана или уничтожена. Второй закон утверждает, что не может быть ни одного процесса, связанного с превращением энергии, без потерь некоторой ее части. Определенное количество энергии в таких превращениях рассеивается в недоступную тепловую энергию, а следовательно, теряется. Отсюда не может быть превращений, к примеру, пищевых веществ в вещество, из которого состоит тело организма, идущих со 100-процентной эффективностью. Таким образом, живые организмы являются преобразователями энергии. И каждый раз, когда происходит превращение энергии, часть ее теряется в виде тепла. В конечном итоге вся энергия, поступающая в биотический круговорот экосистемы, рассеивается в виде тепла. Живые организмы фактически не используют тепло как источник энергии для совершения работы — они используют свет и химическую энергию. Пищевые цепи и сети, трофические уровни. Внутри экосистемы содержащие энергию вещества создаются автотрофными организмами и служат пищей для гетеротрофов. Пищевые связи — это механизмы передачи энергии от одного организма к другому. Типичный пример: животное поедает растения. Это животное, в свою очередь, может быть съедено другим животным. Таким путем может происходить перенос энергии через ряд организмов — каждый последующий питается предыдущим, поставляющим ему сырье и энергию (рис. 12.20).
Рис. 12.20. Биотический круговорот веществ: пищевая цепь (по А. Г. Банникову и др., 1985)
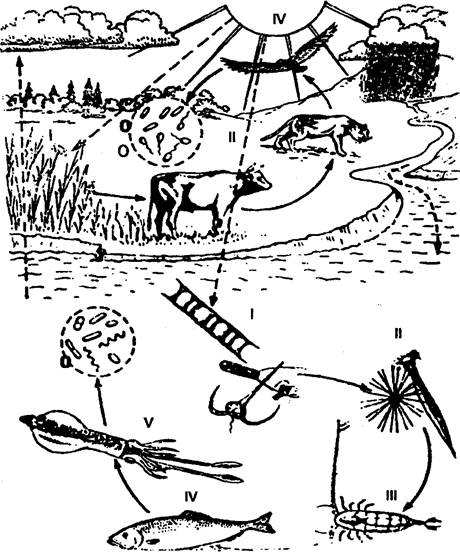
Такая последовательность переноса энергии называется пищевой (трофической) цепью, или цепью питания. Место каждого звена в цепи питания является трофическим уровнем. Первый трофический уровень, как уже было отмечено ранее, занимают автотрофы, или так называемые первичные проду-з центы. Организмы второгого трофического уровня называются первичными консументами, третьего — вторичными консументами и т. д. Обычно различают три типа пищевых цепей. Пищевая цепь хищников начинается с растений и переходит от мелких организмов к организмам все более крупных размеров. На суше пищевые цепи состоят из трех-четырех звеньев. Одна из простейших пищевых цепей имеет вид (см. рис. 12.5): растение ® заяц ® волк продуцент ® травоядное ® плотоядное Широко распространены и такие пищевые цепи: растительный материал (например, нектар) ® муха ® паук ® землеройка ® сова. сок розового куста ® тля ® божья (тлевая) коровка ® ® паук ® насекомоядная птица ® хищная птица. В водных и, в частности, морских экосистемах пищевые цепи хищников, как правило, длиннее, чем в наземных. Широко распространен тип пищевых отношений, представленный на рис. 12.21 и табл. 12.5.
Рис. 12.21. Пищевые цепи в наземной и водной экосистемах: I — продуценты; II — травоядные; III, IV, V — плотоядные; 0 — деструкторы (из Ф. Рамада,1981)
Таблица 12.5
|
||||||
|
|
Последнее изменение этой страницы: 2016-04-20; просмотров: 654; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.156 (0.011 с.) |